
e-ISSN: 2215-3896.
(Enero-Junio, 2022). Vol 56(1)
DOI: https://doi.org/10.15359/rca.56-1.12
Open Acces: www.revistas.una.ac.cr/ambientales
e-Mail: revista.ambientales@una.cr
Licencia: CC BY NC SA 4.0
TECHNICAL NOTE
Diversity and activity patterns of medium and large terrestrial mammals in the Lapa Verde Wildlife Refuge, Heredia, Costa Rica
Diversidad y patrones de actividad de mamíferos terrestres medianos y grandes en el Refugio de Vida Silvestre Lapa Verde, Heredia, Costa Rica
Alejandro José Mattey Trigueros1, David Mattey Trigueros2, Jonathan Navarro Picado3, Lilliana Piedra-Castro4
[Received: June 3, 2021; Accepted: October 14, 2021; Revised: October 31, 2021; Published: January 1, 2022]
Abstract
[Introduction]: Wildlife refuges are crucial to improve connectivity in the context of biological corridors, as they conserve forest patches and consequently neotropical mammal populations. However, in order to devise conservation and management strategies, it is a priority to carry out field studies that validate these efforts and measure their effectiveness over time. [Objective]: To generate baseline information on the population status of medium and large terrestrial mammals and their activity patterns to help the management and protection of the Lapa Verde Wildlife Refuge. [Methods]: We analyzed the camera-trapping methodology by placing 13 camera traps per sampling block, divided into two blocks (1 855 ha) during January-February 2015 and December 2015-January 2016. [Results]: The sampling effort was 1 584 nights, capturing 18 species with the presence of large and medium predators such as Panthera onca, Canis latrans, Leopardus pardalis, and Leopardus wiedii, as well as species of prey such as Silvilagus sp., Dasypus novemcinctus, Dasyprocta punctata, and Cuniculus paca. The species with the highest RAI was D. punctate. The richness was similar between blocks. However, the patterns of circadian activity differed between some species (L. pardalis and D. punctata). [Conclusions]: Species such as jaguars and tapirs, which require large areas of land to settle, use these conservation blocks; this indicates the importance of the conservation of these blocks.
Keywords: biodiversity, predators, prey, protected area management, relative abundance index
Resumen
[Introducción]: Los refugios de vida silvestre son cruciales para mejorar la conectividad en el contexto de corredores biológicos, ya que conservan fragmentos de bosque y consecuentemente poblaciones de mamíferos neotropicales, sin embargo, para poder idear estrategias de conservación y manejo es prioritario realizar estudios de campo que validen estos esfuerzos y midan su efectividad en el tiempo. [Objetivo]: Generar información de línea base sobre el estado poblacional de mamíferos terrestres medianos y grandes y sus patrones de actividad, que contribuyan a realizar un adecuado manejo y protección de este grupo en el Refugio de Vida Silvestre Lapa Verde. [Metodología]: Se utilizó la metodología de foto-trampeo colocando 13 cámaras-trampa por bloque de muestreo, en dos bloques (1 855 ha) durante enero - febrero 2015 y entre diciembre 2015 - enero 2016. [Resultados]: El esfuerzo de muestreo fue de 1 584 noches, registrando18 especies con presencia de grandes y medianos depredadores como Panthera onca, Canis latrans, Leopardus pardalis y Leopardus wiedii, así como especies que sirven de presas como Silvilagus sp., Dasypus novemcinctus, Dasyprocta punctata y Cuniculus paca. La especie con mayor IAR fue D. punctata, la riqueza fue similar entre bloques; sin embargo, los patrones de actividad circadiana sí presentaron diferencias entre algunas especies (L. pardalis y D. punctata). [Conclusiones]: Especies como jaguares y tapires, que requieren grandes extensiones de tierra para asentarse, utilizan estos bloques de conservación, lo que indica la importancia de la conservación de estos bloques.
Palabras clave: biodiversidad; depredadores; índice abundancia relativa; manejo de áreas protegidas; presas.
Worldwide, there are approximately 6,495 described species of mammals (Burgin et al., 2018), of which 253 are described for Costa Rica. They include 115 species of bats, 31 marine and 222 species of terrestrial mammals (Carrillo et al., 2002; Rodríguez-Herrera et al., 2014; Villalobos et al., 2018; Wainwrigth, 2007). Their importance lies in the interaction at the level of ecosystem processes, including pollination, seed dispersion, and predator and prey relationships, which directly influence the structure and composition of vegetation and mediate in the nutrient cycle (Boddicker et al., 2002).
In Costa Rica, there is a necessity for generating information about mammals because some areas of the country lack information about their diversity and conservation status. Specific research in the biological corridors’ areas is scarce but essential to promote effective management actions that result in the reduction of the threats that mammals suffer (Sáenz, 2010). These types of investigations are important to study the influence of connectivity in the study on mammals to validate conservation and management strategies with scientific data. Despite the extensive use of camera traps, it is one of the least used methods to study the effects of connectivity on biodiversity in Latin America (Leija & Mendoza, 2021).
Areas that provide this connectivity for species with ecological importance, such as medium and large mammals, are of importance since these species are highly affected by processes of deforestation, hunting, and isolation that are experienced in the region. They generate pressure on the populations of these species and condition their future survival (Meyer et al. 2019). In addition, these areas represent an important conservation strategy trying to reduce pressure on protected areas and being used as environmental education tools while combining the protection of wildlife with economic activities such as crops or livestock (Zumbado-Morales & Aguero-Valverde, 2020).
For example, Costa Rica shows important conflicts regarding deforestation due to the extension of the agricultural frontier, as well as other conservation threats like illegal logging, which respond to economic, social, cultural, political, and legal causes. This has affected primarily the harvest of all species in primary forests resulting in an impact on landscapes (Programa Estado de la Nación, 2020). In spite of the accelerated rate of deforestation and the high fragmentation of forests that have occurred to date, ambitious projects, like biological corridors, represent an opportunity to mitigate the effects of climate change (Canet-Desanti et al., 2012). One of these important projects is the Mesoamerican Biological Corridor (MBC), launched in 1999. It is a strategy aiming to create a regional corridor that extends from southern Mexico to Panama. This allows the connection between protected areas and remnants of Central American forest (SINAC-MINAE, 2008). Part of this corridor, Lapa Verde Wildlife Refuge (LVWR), is considered a key area for the connection of the MBC since this area has a high value for its biodiversity, so it is an indispensable territory for connectivity between the Caribbean northern zone and the conservation areas in the center of Costa Rica. This allows the country to work as a bridge between North and South America. Thus, it is essential to generate information about the population status of the species that inhabit it (Chassot & Monge, 2002).
This research effort represents the first initiative that Lapa Verde Wildlife Refuge carried out to evaluate the medium and large mammal’s diversity in its forests. The aim of this study is to generate baseline information on the diversity of medium and large terrestrial mammals and their activity patterns in the Lapa Verde Wildlife Refuge, Sarapiquí, Heredia, Costa Rica. The data generated will inform science-driven decision-making and management of mammals.
The Lapa Verde Wildlife Refuge (LVWR) is in the northeast of Costa Rica, Sarapiquí, Heredia Province (Figure 1). It extends over an area of approximately 1 800 ha, of which approximately 1400 are covered with forest. The refuge is divided into two blocks. Block I (B I) with 685 ha, which is in the community of Chilamate at 6 km from the Puerto Viejo town (Figure 2). Block II (B II), with 1 170 ha, is in La Virgen of Sarapiquí, in the communities of California Tico, Río Magdalena, and Pueblo Nuevo (Figure 3).
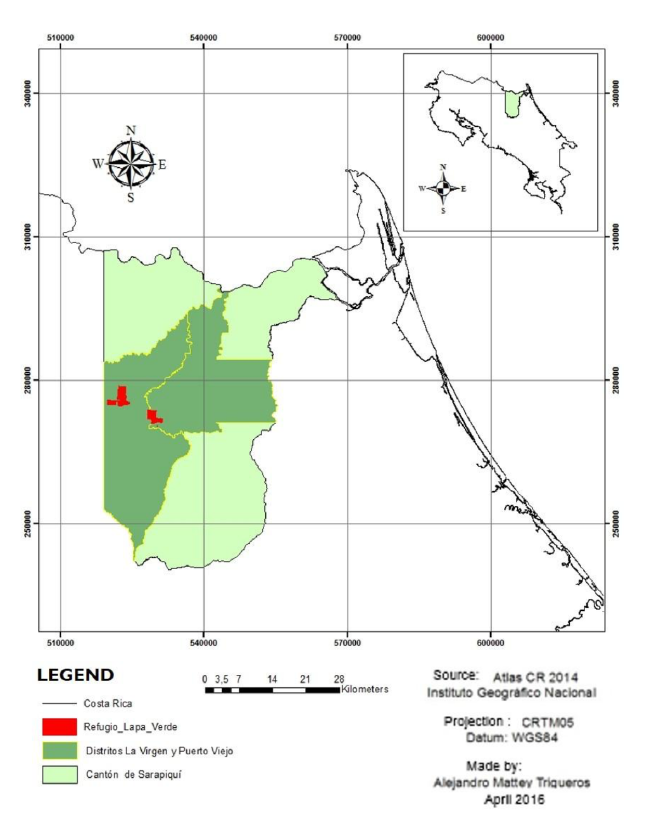
Figure 1. Location of Lapa Verde Wildlife Refuge, La Virgen de Sarapiquí, Heredia, Costa Rica.
Figura 1. Ubicación del Refugio de Vida Silvestre Lapa Verde, La Virgen de Sarapiquí, Heredia, Costa Rica.
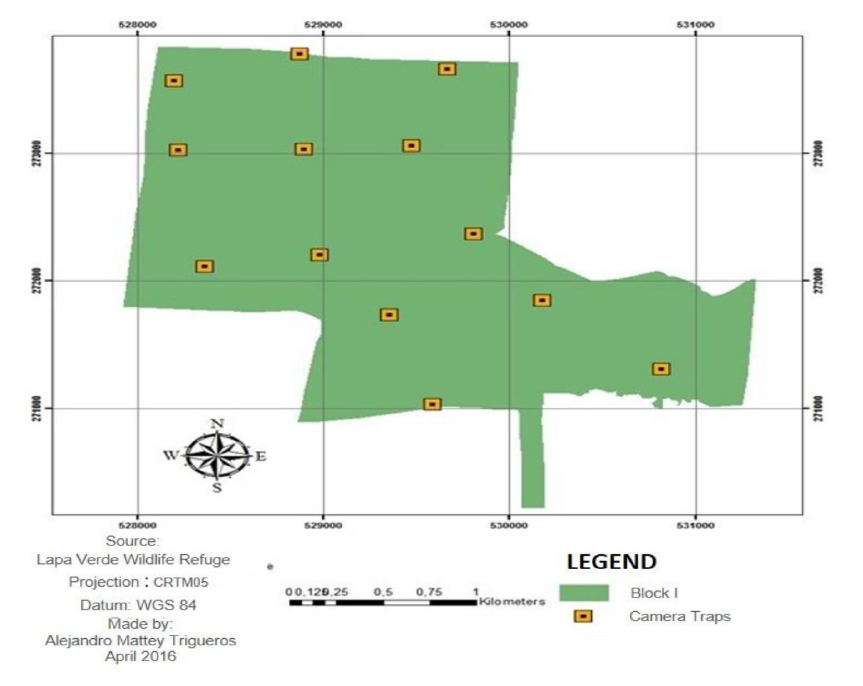
Figure 2. Location of the camera traps in the sampling block I. Lapa Verde Wildlife Refuge, Heredia, Costa Rica.
Figura 2. Ubicación de las cámaras trampa en bloque de muestreo I. Refugio de Vida Silvestre Lapa Verde, Heredia, Costa Rica.
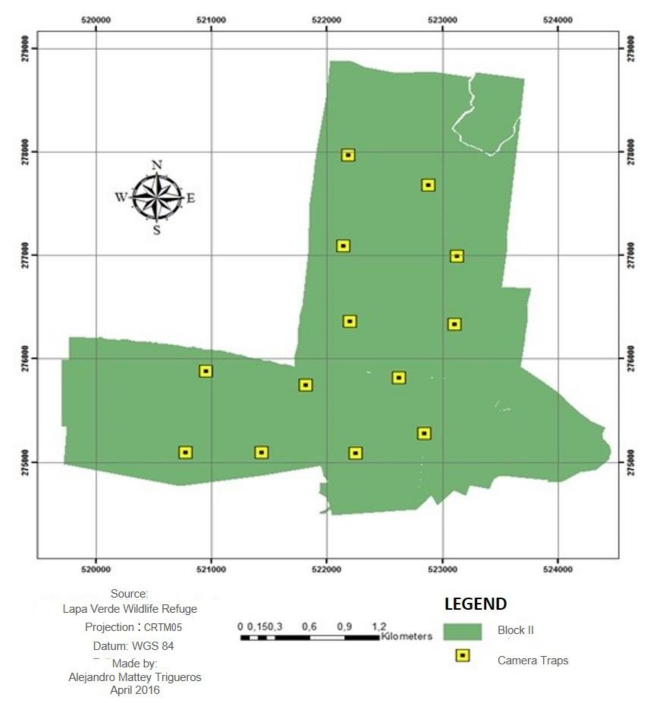
Figure 3. Location of camera traps in block II of sampling. Lapa Verde Wildlife Refuge, Heredia, Costa Rica.
Figura 3. Ubicación de las cámaras trampa en bloque de muestreo II. Refugio de Vida Silvestre Lapa Verde, Heredia, Costa Rica.
According to the Holdridge’s life zones classification, the predominant life zone in the area is Very Humid Tropical Forest (bmh-T). It is characterized by high precipitation with averages ranging from 3 000 to 5 400 mm, and with an average temperature of 26.1 °C, ranging from 20.7 °C to 32.5 °C (Holdridge & Tosi, 1967).
The main productive activities that take place in the surroundings of the LVWR are livestock and expansive pineapple plantations. Other activities present in lower intensity are crops of basic grains, tubers, heart of palm plantations, and some monoculture plantations of exotic timber species such as Melina (Gmelina arborea) and Teak (Tectona grandis) (Chassot & Monge, 2002).
In this study, we used the camera-trapping methodology (Karanth & Nichols, 1998). According to Monroy et al. (2010), it is one of the most effective techniques for estimating the abundance of wild species and achieving an evaluation of the population’s status for their conservation. This study was carried out in two phases to cover the two conservation blocks managed by the Lapa Verde Wildlife Refuge. The first phase of data collection was carried out during the months of January and February 2015 in the Sampling Block I (Chilamate); the second phase was developed during the months of December 2015 and January 2016 in Block II (Pueblo Nuevo).
In each sampling block (Figure 2 and Figure 3) 13 camera traps were installed (Bushnell Trophy Camera, Model 119537 and Moultrie, model MCG-12596). The equipment was attached to trees and secured with safety chains at 50 cm from the ground, in order to avoid camera extraction and loss of data due to equipment theft. Each device was installed at an average distance of 750 m of each other and placed on passages where animal tracks and signs were present (Díaz-Pulido & Payán-Garrido, 2012). The surroundings of each camera were clear areas without dense understory to allow the capture of better images.
The cameras were programmed to remain active for 24 h (necessary to calculate night sampling) and olfactory attractants were used in each one, consisting of stakes with pieces of cotton cloth dipped completely with Obsession perfume of the Kalvin Klein brand (Monroy et al., 2010; Gontier & Castañeda, 2013; Díaz-Pulido & Payán-Garrido, 2012). The cameras were checked every 15 days to verify their proper functioning, change batteries and memory cards if necessary.
Data Analysis. The identification of the species photographed was done using field guides (Wainwrigth, 2007, Reid 2009). Data were analyzed using EstimateS statistical software, (Díaz & Payan, 2012) and the statistical program R version 3.5.1. Relative Abundance Index (RAI) was carried out for independent photographs, using Equation 1. To estimate abundance more accurately, we used the technique by Monroy et al. (2010).
RAI= events*100 camera trap nights/sampling effort (E.1)
Where:
RAI: relative abundance index for species ‘a’
Events: number of independent records per species
100 camera trap nights: unit of standardization to compare data with other studies
Sampling effort: total amount of nights that the camera trap stations were working
To determine the patterns of circadian activity, a record of the occurrence of the species was made, analyzing the frequency (Number of photos in every interval of time) of activity of each to calculate the time of day they were more active. Only the species with a minimum of 11 independent photographic records and had the time set in the photograph were taken into account. This number of records has been considered as the minimum necessary to describe the pattern of circadian activity of a species (Maffei et al., 2002). The data obtained were ordered in four-hour intervals to define and perform the analysis of the circadian activity pattern by species in these ranges and contrast between the results obtained between both blocks.
A sampling effort of 795 camera nights (nights installed on the field per amount of cameras) for Block I and 789 in Block II was made for a total of 1,584 camera nights. A total of 18 species distributed in 12 families were captured, of which 15 species are shared between both sampling blocks (Table 1). Only 6 species, Canis latrans, Panthera onca, Galictis vittata, Silvilagus gabbi, Procyon lotor, and Tapirus bairdii were not shared, corresponding to 33.3 % of the recorded species. Among the families that were registered, Felidae is the most representative with 4 species: Panthera onca, Leopardus pardalis, Leopardus wiedii and Puma yagouaroundi. In addition, 6 trophic guilds were found, and the presence in the study area of large and medium predators such as P. onca, C. latrans, L. pardalis, and L. wiedii should be highlighted.
In addition, there are herbivorous and frugivorous species in the area, for example, Silvilagus sp., Tapirus bairdii, Dasypus novemcinctus, Dasyprocta punctata and Cuniculus paca (Table 1). On the other hand, the conservation status of the captured species is important to take in consideration. For example, P. onca and L. wiedii that are near threatened and M. temama which does not have enough data to generate an accurate conservation status (Table 1).
Table 1. Medium and large mammal species recorded in both sampling blocks (I and II), with their respective trophic guild and conservation status according to IUCN. Lapa Verde Wildlife Refuge, Heredia, Costa Rica.
Cuadro 1. Especies de mamíferos medianos y grandes registrados en ambos bloques de muestreo (I y II), con su respectivo gremio trófico y estatus de conservación según la UICN. Refugio de Vida Silvestre Lapa Verde, Heredia, Costa Rica.
|
Family |
Specie |
Trophic Guild |
Local name |
UICN |
|
Canidae |
Canis latrans |
C |
Coyote |
LC |
|
Cuniculidae |
Cuniculus paca |
G, F |
Tepezcuintle |
LC |
|
Dasyproctidae |
Dasyprocta punctata |
G, F |
Guatusa |
LC |
|
Dasypodidae |
Dasypus novemcinctus |
I |
Armadillo |
LC |
|
Mustelidae |
Eira barbara |
O |
Tolomuco |
LC |
|
Galictis vittata |
O |
Grisón |
LC |
|
|
Felidae |
Leopardus pardalis |
C |
Manigordo |
LC |
|
Leopardus wiedii |
C |
Caucel |
NT |
|
|
Puma yagouaroundi |
C |
Jaguaroundi |
LC |
|
|
Panthera onca |
C |
Jaguar |
NT |
|
|
Procyonidae |
Nasua narica |
O |
Pizote |
LC |
|
Procyon lotor |
O |
Mapache |
LC |
|
|
Cervidae |
Odocoileus virginianus |
H |
Venado cola blanca |
LC |
|
Mazama temama |
H |
Cabro de monte |
DD |
|
|
Tayassuidae |
Pecari tajacu |
H |
Saíno |
LC |
|
Leporidae |
Silvilagus gabbi |
H |
Conejo de monte |
NA |
|
Myrmecophagidae |
Tamandua mexicana |
I |
Oso hormiguero |
LC |
|
Tapiridae |
Tapirus bardii |
H |
Danta |
LC |
C = carnivorous, G = granivorous, F = frugivorous, I = insectivorous, O = omnivorous, H = herbivore. In addition, its conservation status according to IUCN with acronym in English, LC = Least concern, NT = Near threatened, DD = deficient data and NA = not applicable since no identification was made at the species level.
The species with the highest RAI for both blocks was D. punctata (Block I: 80.50 and Block II: 63.37) and showed similar values and number of catches at the two sampling sites. N. narica showed the second highest value of RAI in Block II (63.37); however, in Block I it was only registered on one occasion obtaining a very low RAI (1.25). Some species, such as D. novemcinctus (Block I, RAI = 22.64, Block II, RAI = 30.42) and L. pardalis (Block I, RAI = 16. 35, Block II, RAI = 12.67), obtained similar records and therefore a very similar RAI for both sampling blocks.
There were species that showed a single record, such as P. onca, which registered only in Block I and G. vittata that failed to be registered in Block I and was recorded in a single occasion in Block II. In the case of T. bardii, it showed only two records in Block II, while P. yagouarondi. L. wiedii who obtained a few records in one of the blocks and other only one record in the other block. It differs from this pattern C. latrans that despite presenting 13 records in Block I (RAI = 16.35) it was not registered for block II. For the other species, differences of records were found between the two sampling blocks, such as P. tajacu, O. virginianus, C. paca, T. mexicana, M. mazama and E. barbara (Table 2).
Table 2. Number of records and Relative abundance (RAI) of medium and large mammals´ species recorded by sampling block in the study area.
Cuadro 2. Número de registros y Abundancia relativa (IAR) de especies medianas y grandes de mamíferos por bloque de muestreo.
|
Species |
Records Block I |
RAI Block I |
Records Block II |
RAI Block II |
|
Canis latrans |
13 |
16.35 |
0 |
0.00 |
|
Cuniculus paca |
5 |
3.77 |
7 |
8.87 |
|
Dasyprocta punctata |
90 |
80.50 |
50 |
63.37 |
|
Dasypus novemcinctus |
20 |
22.64 |
24 |
30.42 |
|
Eira barbara |
15 |
16.35 |
5 |
6.34 |
|
Galictis vittata |
0 |
0.00 |
1 |
1.27 |
|
Leopardus pardalis |
19 |
16.35 |
10 |
12.67 |
|
Leopardus wiedii |
1 |
1.25 |
4 |
5.07 |
|
Puma yagouaroundi |
4 |
5.03 |
1 |
1.27 |
|
Panthera onca |
1 |
1.03 |
0 |
0.00 |
|
Nasua Narica |
10 |
1.25 |
50 |
63.37 |
|
Procyon lotor |
2 |
2.51 |
0 |
0.00 |
|
Odocoileus virginianus |
2 |
2.51 |
14 |
17.74 |
|
Mazama americana |
0 |
0.00 |
5 |
6.34 |
|
Pecari tajacu |
8 |
10.06 |
19 |
24.08 |
|
Silvilagus gabbi |
4 |
5.03 |
0 |
0.00 |
|
Tamandua mexicana |
4 |
5.03 |
7 |
8.87 |
|
Tapirus bairdii |
0 |
0.00 |
2 |
2.54 |
3.3 Circadian activity patterns
The comparison between patterns of circadian activity presented in Figure 4 and Figure 5 shows patterns among the species that were shared between the study blocks. Since 3 species presented different patterns in terms of their peak activity, like L. pardalis, with the highest nocturnal activity in Block I, while Block II, the highest number of records were obtained in the morning and mid-day. D. novemcinctus, meanwhile, was recorded throughout the day in the Block I while in Block II catches were focused on night hours, showing a difference of activity of the same species between the two blocks. Other species showing more activity during the day were P. tajacu, N. narica, and D. punctate,
In the case of L. pardalis, the exception should be made that its activity pattern was included in the analysis despite having 10 catches (less than recommended to perform an activity pattern as mentioned in the Methods section). This was made in order to obtain data and patterns that allow identifying specific conditions of the sampling site to propose management and protection actions for this species. The exception was made considering that the species is a common predator in the study area.
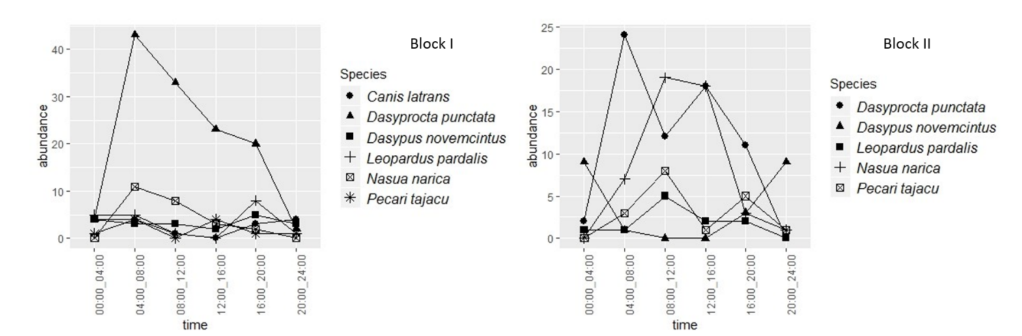
Figure 4. Circadian activity patterns by species in Block l and Block II of the study area.
Figura 4. Patrones de actividad circadiana por especies en el Bloque l y Bloque II de muestreo.
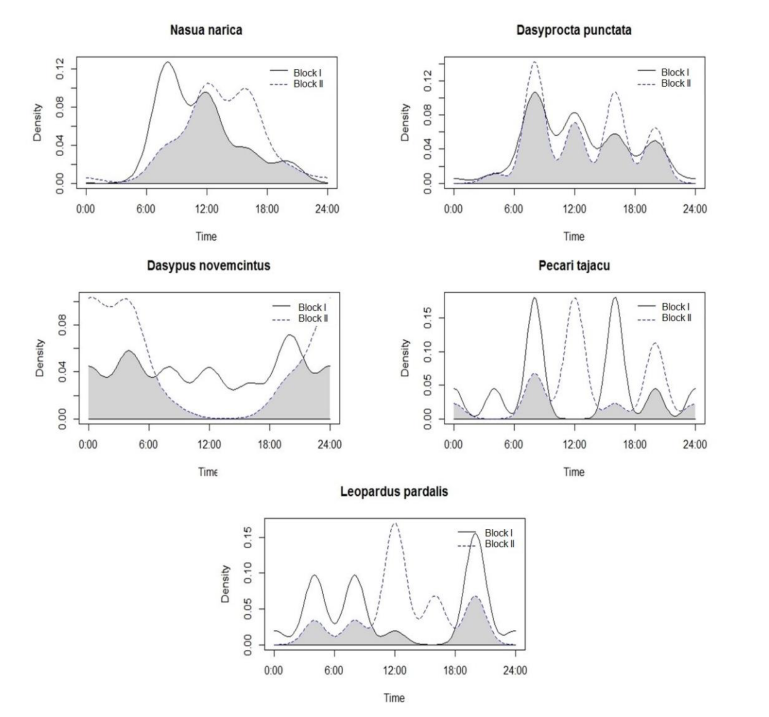
Figure 5. Activity patterns by 5 species in sampling blocks I and II of the study site.
Figura 5. Patrones de actividad por especie en ambos bloques de muestreo.
The number of species recorded in this study was similar compared to studies carried out in the region, in which the majority of medium and large mammal species are shared. For example, Salom et al. (2015) with a sampling effort of 3 640 nights (higher than this study) recorded 22 species, compared to the 18 captured in our sampling area. In another study, Arroyo-Arce et al. (2017), also in the Caribbean lowlands of Costa Rica, had a sampling effort quite similar (1 643 nights) to the present study and found 11 species with carnivores as the most common group, including three species of felines. Something important to highlight is the fact that L. pardalis was the feline species with the highest number of catches, this coincides with different studies in different areas of the country (Bustamante, 2008; Corrales-Gutiérrez et al. 2012; Salom et al., 2015; Sáenz, 2010) and is probably in response to the ability of this species to colonize both disturbed and dense forests sites (Cruz-Rodríguez, 2012; Pérez-Irineo & Santos-Moreno, 2014).
The Wildlife Refuge allows the establishment of populations of umbrella species with different habitat selection criteria, such as P. onca, which is a species more related to dense forests and sites slightly altered in contrast to N. narica associated with more open areas, with lower forest cover density (Foster & Vaughan, 2002; Pacheco et al., 2006). In addition, it should be noted that of the 18 species found throughout the study, 5 have a carnivorous trophic guild, which indirectly indicates the quality and condition of the study area because the presence of predators and top predators such as Jaguars are good indicators of prey populations in the trophic chain due to predator-prey interactions. (Crooks et al., 2011; Mora, 2017). Besides, there are species that serve as prey for large and medium predators and perform important ecological functions such as seed dispersal. This allows maintaining the life cycle of many plants and their roles in the ecosystem, for example, Silvilagus sp., Tapirus bairdii, Dasypus novemcinctus, Dasyprocta punctata, and Cuniculus paca (Mittelman et al., 2020; Wainwrigth, 2007).
The activity patterns recorded in this study show that data regarding the behavior of the same species varies between sampling blocks. These differences can be attributed to three main causes related to the degree of anthropogenic disturbance present in the study site.
1) In Block II there is a constant movement of machinery to attend the pineapple production farm. Considering the block is immersed in a matrix of pineapple monoculture plantations, this could generate effects on the activity of the species due to noise pollution and vulnerability to roadkill. This may represent a marked difference with respect to Block I in which there is no access to machinery and can only be accessed through trails.
2) Of the different stages of ecological succession between the sampling sites, Block II has large fragments of natural regeneration in early succession stages and is currently used by timber plantations. Block I did not present a disturbance in terms of timber exploitation and is mainly covered by forest in more advanced succession stages than Block II (Rojas-Durán, 2010). This could influence the difference in the patterns of circadian activity between sampling blocks (Acevedo-Charry & Aide, 2019; Paviolo et al. 2009).
3) Poaching can be one of the differences between the degree of conservation of both sampling blocks. As a personal observation, it is noted that complaints of hunting in Block II were more frequent than in Block I. The presence of poaching is crucial to understand data of wealth and circadian activity patterns as highlighted by Di Bitteli et al. (2008) because this activity can affect the hunting of target species and their predators, affecting, in turn, the ecosystem dynamics of the studied area.
These differences between sampling sites could allude to the differences between the blocks’ samplings explained previously, but further research should be conducted for better understanding of the causes of the differences found in this study. The behavior of the species studied will vary according to environmental pressures, both for predatory species and for predated species (Monterroso et al., 2013), which increases the importance of studies that generate the first approach to identify the pressures and possible management actions that can be used in a given area.
The evidence showing that species such jaguars and tapirs, which require large areas of land to settle, use these conservation blocks indicates the importance of the conservation of these blocks, the expanse of conservation lands, and the improvement of their connectivity to provide habitat for these important species.
The information of this study has importance as a baseline for future studies that we consider to be focus on long-term projects and expand the studies to private and national protected areas in the region to have a clear result about the connectivity in this study area. The other important aspect to highlight is that this study was carried out in a period of 4 months which corresponds to the dry season in Costa Rica, so this research represents a baseline on the diversity of mammals in the refuge and some aspects of their activity. Therefore, it is necessary to expand with other studies that consider both climatic seasons in Costa Rica.
An important component to take into account for future studies is the application of occupancy modeling to explain, in a better way, the use of the landscape of medium and large terrestrial mammals.
6. Ethics and conflict of interest
The authors declare that they have fully complied with all pertinent ethical and legal requirements, both during the study and in the production of the manuscript; that there are no conflicts of interest of any kind; that all financial sources are fully and clearly mentioned in the acknowledgments section; and that they fully agree with the final edited version of the article.
To the Ecovida Foundation for the donation of the camera traps. To the Journal and anonymous reviewers for their comments on the latest version of the manuscript.
Acevedo-Charry, O., & Aide, T. M. (2019). Recovery of amphibian, reptile, bird and mammal diversity during secondary forest succession in the tropics. Oikos, 128(8), 1065-1078. https://doi.org/10.1111/oik.06252
Arroyo-Arce, S., Thomson, I., Fernández, C., & Salom-Pérez, R. (2017). Relative abundance and activity patterns of terrestrial mammals in Pacuare Nature Reserve, Costa Rica. Cuadernos de Investigación UNED, 9(1), 15-21. https://doi.org/10.22458/urj.v9i1.1673
Boddicker, M., Rodriguez, J. J., & Amanzo, J. (2002). Indices for assessment and monitoring of large mammals within an adaptive management framework. Environmental Monitoring and Assessment, 76(1), 105-123.Burgin, CJ, Colella, JP, Kahn, PL y Upham, NS (2018). ¿How many species of mammals there are? Journal of Mammalogy, 99 (1), 1-14.
https://doi.org/10.1093/jmammal/gyx147
Burgin, C. J., Colella, J. P., Kahn, P. L., & Upham, N. S. (2018). How many species of mammals are there?. Journal of Mammalogy, 99(1), 1-14.
Bustamante, A. (2008). Densidad y uso de hábitat por los ocelotes (Leopardus pardalis), pumas (Puma concolor) y jaguares (Panthera onca) en la parte sureste del área de amortiguamiento del Parque Nacional Corcovado, Península de Osa. Costa Rica. Tesis de Maestría. Instituto Internacional en Conservación y Manejo de Vida Silvestre.
Canet-Desanti, L., Herrera, B., & Finegan, B. (2012). Efectividad de manejo en corredores biológicos: el caso de Costa Rica. Revista Parques, 2, 1-10.
Carrillo, E. Wong, G & Sáenz, J. (2002). Mamíferos de Costa Rica. 2 ed. Santo Domingo de Heredia. Instituto Nacional de Biodiversidad (INBIO). 2002
Chasot, O. & Monge, G. (2002). Ficha Técnica Corredor Biológico San Juan- La Selva. Costa Rica.
Corrales-Gutiérrez D., R. Salom-Pérez, J. Carazo-Salazar & D. Araya. (2012). Evaluación inicial de la Unidad de Conservación del Jaguar (Panthera onca) Parque Nacional Braulio Carrillo, Costa Rica. Informe técnico. PANTHERA, Costa Rica.
Crooks, K., Burdett, C., Theobald, D., Rondinini, C., &Boitani, L. (2011). Global patterns of fragmentation and connectivity of mammalian carnivore habitat. Phil. Trans. R. Soc. B., 366, 2642–2651. https://doi.org/10.1098/rstb.2011.0120
Cruz-Rodríguez C. A. (2012). Ecología espacial del ocelote (Leopardus pardalis) en un paisaje fragmentado del Municipio de Colosó (sucre), Caribe colombiano. Mastozool. Neotrop 19 (2), 367-383.
Di Bitteli, M. S., Paviolo A., Ferrari, C. A., De Angelo C. y Di Blanco, Y. (2008). Differential responsees to huntint in two sympatric species of brocket deer (Mazama Americana and M. nana). Biotropica, 40(5), 636-645. https://doi.org/10.1111/j.1744-7429.2008.00413.x
Díaz-Pulido, A. y E. Payán-Garrido. (2012). Manual de foto-trampeo: una herramienta de investigación para la conservación de la biodiversidad en Colombia. Instituto de Investigaciones de Recursos Biológicos Alexander von Humboldt y Panthera Colombia.
Foster C. & Vaughan, C. (2002). Home range, habitat use, and activity of Baird’s Tapir in Costa Rica. Biotropical, 34 (3), 423-437. https://doi.org/10.1111/j.1744-7429.2002.tb00556.x
Gonthier, D. & J. Castañeda, F. E. (2013). Large- and medium-sized mammal survey using camera traps in the Sikre River in the Río Plátano Biosphere Reserve, Honduras. Tropical Conservation Science, . 6(4), 584-591. https://doi.org/10.1177/194008291300600409
Holdridge, L. R., & Tosi, J. A. (1967). Life zone ecology. Rev.ed. ed. San Jose, Costa Rica.
Karanth, K. U. & J. D. Nichols. (1998). Estimation of tiger densities in India using photographic captures and recaptures. Ecology, 79, 2852-2862. https://doi.org/10.1890/0012-9658(1998)079[2852:EOTDII]2.0.CO;2
Leija, E. G. & M. E. Mendoza. (2021). Estudios de conectividad del paisaje en América Latina: retos de la investigación. Madera y Bosques, 27(1), e2712032.
https://doi.org/10.21829/myb.2021.2712032
Maffei, L., E. Cuellar & J. Noss. (2002). Uso de trampas-cámara para la evaluación de mamíferos en el ecotono Chaco-Chiquitanía. Revista boliviana de ecología y conservación ambiental, 11, 55-65.
Meyer, N. F. V., R. Moreno, C. Sutherland, J. A. de la Torre, H. J. Esser, C. A. Jordan, M. Olmos, J. Ortega, R. Reina-Hurtado, S. Valdes & P. A. Jansen. (2019). Effectiveness of Panama as an intercontinental land bridge for large mammals. Conservation Biology, 34(1), 1-13.
https://doi.org/10.1111/cobi.13384
Mittelman, P., Kreischer, C., Pires, A. S., & Fernandez, F. A. (2020). Agouti reintroduction recovers seed dispersal of a large-seeded tropical tree. Biotropica, 52(4), 766-774. https://doi.org/10.1111/btp.12788
Monroy, O. Zarco, M. Rodríguez, C. Soria. L. Urios, V. (2010). Estudio Foto-trampeo de mamíferos en la Sierra Nanchititla, México: abundancia relativa y patrón de actividad. Rev. Biol. Trop. 59 (1). https://doi.org/10.15517/rbt.v59i1.3206
Monterroso, P., P. C. Alves & P. Ferreras. (2013). Catch me if you can: diel activity patterns of mammalian prey and predators. Ethology 119, 1044–1056. https://doi.org/10.1111/eth.12156
Mora, F. (2017). Nation-wide indicators of ecological integrity in Mexico: The status of mammalian apex-predators and their habitat. Ecological Indicators, 82, 94-105.
https://doi.org/10.1016/j.ecolind.2017.06.030
Pacheco J., G. Ceballos, G. Daily, P. Ehrlich, G. Suzán, B. Rodríguez & E. Marcé. (2006). Diversidad, historia natural y conservación de los mamíferos de San Vito Coto Brus, Costa Rica. Rev. Biol. Trop. 54 (1), 219-240. https://doi.org/10.15517/rbt.v54i1.13998
Paviolo, A., Di Blanco, Y. E., De Angelo, C. D., y Di Bitetti M. S. (2009). Protection affects the abundance and activity patterns of pumas in the Atlantic forest. J. Mammal 90 (4), 926-934. https://doi.org/10.1644/08-MAMM-A-128.1
Pérez-Irineo, G. & A. Santos-Moreno. (2014). Density, distribution, and activity of the ocelot Leopardus pardalis (Carnivora: Felidae) in Southeast Mexican rainforests. Rev. Biol. Trop. 62 (4), 1421-1432. https://doi.org/10.15517/rbt.v62i4.12941
Programa de La Nación, P. E. (2020). Estado de la nación en desarrollo humano sostenible. Séptimo Informe. San José, Costa Rica.
Reid, F.A. (2009). A field guide to the mammals of Central America & Southeast Mexico. Oxford, U.K.: Oxfort University Press.
Rodríguez-Herrera, B., Ramírez-Fernández, J. D., Villalobos-Chaves, D., & Sánchez, R. (2014). Actualización de la lista de especies de mamíferos vivientes de Costa Rica. Mastozoología neotropical, 21(2), 275-289.
Rojas-Durán, Y. (2010). Plan de Manejo del Refugio Lapa Verde. Heredia, CR, Ecovida S.A. 96 p.
Sáenz, C. (2010). Ensamble de mamíferos medianos y grandes en un sector de la Reserva Forestal Río Pacuare y sus cercanías (Reserva Indígena NairiAwari y Parque Nacional Barbilla, Costa Rica. Tesis presentada para optar al grado de Magister Scientiae en Conservación y Manejo de Vida Silvestre. Universidad Nacional. Heredia. Noviembre 2010, Costa Rica.
Salom-Pérez R., D. Araya-Gamboa, D., Corrales Gutiérrez, J. Carazo-Salazar, E. Urbina, D. Espinoza-Muñoz, E. Pomareda, C. Aguilar, S. Arroyo-Arce, C. Wultsch, A. Caragiulo, L. Petracca, H. Robinson, N. Robinson, S. Soto, G. Gutiérrez, M. Spínola, M. Chávez & H. Quigley. (2015). Programa de monitoreo de Mamíferos Medianos y Grandes en el Sub-corredor Biológico Barbilla-Destierro/paso del jaguar informe final. Corporación Panthera.
SINAC-MINAE. (2008). Diseño, oficialización y consolidación de corredores biológicos en Costa Rica /– 1 ed.- San José, C.R.: Comité de Apoyo a los corredores biológicos, 2008.
Villalobos-Chaves, D., González-Quirós, A., Lara-Hernández, L. y Rodríguez-Herrera, B. (2018). Notes on the geographic range and distribution of two free-tailed bat species (Chiroptera, Molossidae) in Costa Rica. Check List., 14, 805. https://doi.org/10.15560/14.5.805
Wainwrigth, M. (2007). The Mammals of Costa Rica. 1 ed. Cornell University Press. Ithaca and London.
Zumbado-Morales, F. & Agüero –Valverde. (2020). Connectivity, ecosystem services and Nature-based solutions in land-use planning in Costa Rica. En: Guidelines for conserving connectivity through ecological networks and corridors. UICN. Switzerland. (98-99).
1 Escuela de Ciencias Exactas y Naturales, Universidad Nacional a Distancia, Costa Rica. alemattey@hotmail.com;
https://orcid.org/0000-0001-7787-9197
2 Escuela de Ciencias Biológicas, Universidad Nacional, Costa Rica. davidmattey@gmail.com; https://orcid.org/0000-0002-6526-9591
3 Escuela de Ciencias Biológicas, Universidad Nacional, Costa Rica. jonathan.navarro.picado@una.cr; https://orcid.org/0000-0003-3380-9546
4 Laboratorio de Recursos Naturales y Vida Silvestre (LARNAVISI), Escuela de Ciencias Biológicas, Universidad Nacional, Costa Rica. lilliana.piedra.castro@una.cr; https://orcid.org/0000-0003-4878-1531



Escuela de Ciencias Ambientales,
Universidad Nacional, Campus Omar Dengo
Apartado postal: 86-3000. Heredia, Costa Rica
Teléfono: +506 2277-3688
Correo electrónico revista.ambientales@una.ac.cr