
e-ISSN: 2215-3896.
(Enero-Junio, 2024). Vol 58(1)
DOI: https://doi.org/10.15359/rca.58-1.7
Open Acces: www.revistas.una.ac.cr/ambientales
e-Mail: revista.ambientales@una.cr
Licencia: CC BY NC SA 4.0
Modelado de idoneidad ambiental e implicaciones del cambio climático en la distribución de Pseudotsuga menziesii (MIRB.) Franco para México
Environmental suitability modeling and implications of climate change on the distribution of Pseudotsuga menziesii (MIRB.) Franco for Mexico
Rodolfo Rodríguez-Ruiz1, Alejandro Juárez-Agis2, Silberio García-Sánchez3, Branly Olivier-Salome4, Enrique Ortiz-Bermúdez5, Rocío Ramírez-Barrios6
[Recibido: 29 de junio 2023, Aceptado: 21 de agosto 2023, Corregido: 20 de septiembre 2023, Publicado: 7 de noviembre 2023]
Resumen
[Introducción] Pseudotsuga menziesii (Mirb.) Franco es una especie forestal de gran importancia por sus servicios ecosistémicos y su comercialización. Actualmente, se ha documentado que el cambio climático está afectando las poblaciones del norte del continente; sin embargo, no se ha evaluado cómo afectaría esto a la distribución de la especie en el futuro. [Objetivo] El objetivo de este trabajo fue modelar la idoneidad ambiental y la distribución potencial de P. menziesii en 3 periodos (2040, 2060 y 2080). [Metodología] Se estimó la idoneidad y distribución potencial, con base en registros y variables bioclimáticas, con ayuda de MaxEnt; se realizaron transferencias a 2 escenarios de emisiones de carbono en 3 periodos; por último, se evaluó la pérdida de extensión y disminución de idoneidad ambiental en porcentaje. [Resultados] P. menziesii tiene una distribución potencial actual de 164 110 km2, las zonas con mayor idoneidad ambiental (> 0.5) se ubicaron en el norte del país, específicamente en los estados del norte. No obstante, bajo los escenarios de cambio climático, estas áreas sufrieron mayor disminución de la idoneidad ambiental (> 50 %) y se redujo su extensión entre 50.7 % y 92.6 %; en contraste, las poblaciones más resilientes contra el cambio climático fueron las del sur. [Conclusiones] Pseudotsuga menziesii se ve favorecida por las bajas temperaturas, debido a esto, se espera un decrecimiento drástico en su distribución potencial. Aunada a ello, la idoneidad ambiental bajará a más del 50 % en las regiones norte de la nación. Con una pérdida drástica de superficie, la prevalencia de la especie pudiera encontrarse en un punto de declive.
Palabras clave: distribución potencial; calentamiento global; MaxEnt; Pinaceae.
Abstract
[Introduction] Pseudotsuga menziesii (Mirb.) Franco is a forest species of great importance for its ecosystem services and commercialization. Currently, it has been documented that climate change is affecting populations in the north of the continent; Nevertheless, how these changes will affect the distribution of the species in the future has not been assessed. [Objective] The objective of this work was to model the environmental suitability and potential distribution of P. menziesii in 3 time periods (2040, 2060 and 2080). [Methodology] Environmental suitability and potential distribution were estimated based on records and bioclimatic variables with the help of MaxEnt; transfers were made to 2 scenarios of carbon emissions in 3 time periods; finally, the loss of extension and decrease of environmental suitability were evaluated as a percentage. [Results] P. menziesii has a current potential distribution of 164 110 km2, the areas with the highest environmental suitability (> 0.5) were located in the north of the country, specifically in the states of north. However, under the climate change scenarios, these zones suffered a greater decrease in their environmental suitability of > 50 %, and their extension was reduced between 50.7 % and 92.6 %, in contrast, the most resilient populations to climate change were those in the south. [Conclusions] Pseudotsuga menziesii is favored by low temperatures, due to this, a drastic reduction in its potential distribution is expected, coupled with this, environmental suitability will drop to more than 50 % in the northern regions of the country. With a drastic loss of area, the prevalence of the species could be at a point of decline.
Keywords: global warming; MaxEnt; Pinaceae; potential distribution.
Pseudotsuga menziesii (Mirb.) Franco es una conífera de importancia económica en México, ya que se utiliza con fines maderables y para plantaciones de árboles de Navidad (Álvarez-Moctezuma et al., 2007). De igual manera, esta especie presenta una alta trascendencia ecológica y es considerada un eficiente reservorio de carbono (García et al., 2021).
Se cree que P. menziesii es tanto abundante como fundamental en el oeste de EE. UU. y Canadá (Álvarez-Moctezuma et al., 2007), sin embargo, en México es escasa y sus poblaciones se encuentran fragmentadas, por ello, está bajo protección especial en la NOM-059-SEMARNAT-2010 (DOF, 2019).
La distribución de P. menziesii para México está conformada por 29 poblaciones dispersas y aisladas, repartidas entre el sur, centro y norte del país (Hernández et al., 2005). Las poblaciones del norte mexicano son las más representativas, en contraste con las del centro y sur, que son pequeños rodales (Ríos et al., 2010).
En P. menziesii se ha documentado que el efecto de las variaciones del clima y la fragmentación puede mermar el número de individuos, el aumento de la temperatura otoñal afecta la resistencia de las poblaciones a las temperaturas invernales (Sheel Bansal et al., 2015). Así, las condiciones climáticas y de los sitios pueden afectar negativamente variables como germinación, producción de semillas, peso de semilla y supervivencia (Juárez-Agis et al., 2006). De igual manera, se ha registrado que las poblaciones de climas más cálidos y secos o con inviernos relativamente fríos y veranos áridos tienen una mayor resistencia a la sequía; con esto, pueden estar adaptadas para hacer frente al panorama que se espera en el futuro (Bansal et al., 2015).
El impacto del cambio climático en los bosques se analiza desde la década de 1990 y ha sido uno de los temas más discutidos por diversos autores (Ledig et al., 2010; Monterroso-Rivas et al., 2013; Peterson et al., 2001; Romero-Sánchez, 2016); así, las alteraciones que pudieran ocurrir en los tipos de vegetación tendrán como consecuencia un aumento o una disminución en su cobertura. Esto resultará en una nueva distribución espacial de las especies; además, habrá implicaciones en la abundancia de algunas, entre las que destacan las más sensibles o vulnerables (Peterson et al., 2001).
Las modificaciones en los patrones de distribución han surgido a lo largo de la historia de la Tierra, sin embargo, en la última década, los efectos negativos han sido impulsados, en gran medida, por el impacto de actividades antrópicas (Ledig et al., 2010; Monterroso-Rivas et al., 2013). De acuerdo con Romero-Sánchez (2016, p.1): “No hay certeza de la magnitud, frecuencia e interacciones que el cambio climático ocasionará en el futuro, por lo que el uso de escenarios climáticos es la forma más cercana para estimar los efectos del calentamiento global en ecosistemas terrestres” y conocer las secuelas que el clima tendrá en las especies.
Actualmente, existen modelos globales de cambio climático en los que cada línea supone un escenario diferente a futuro: alentador, conservador y pesimista. Se asume cómo podría evolucionar el clima en un futuro en el cual se tomarán o no acciones políticas, se usarán combustibles fósiles, habrá crecimiento de la población y desarrollo económico, entre otros (Escoto et al., 2017).
Estos modelos globales, en conjunto con los algoritmos de modelado como BIOCLIM y GARP (Plasencia-Vázquez, 2022) o MaxEnt (Phillips et al., 2017), han sido utilizados para evaluar el impacto en la distribución y permanencia de diversos grupos de especies ante el cambio climático, a partir de variables climáticas y registros de presencia (Ledig et al., 2010; Monterroso-Rivas et al., 2013). Asimismo, han ayudado a conocer el efecto del cambio climático en especies forestales (Guitérrez y Trejo, 2014).
Estas técnicas han permitido generar modelos de idoneidad ambiental, los cuales representan valores continuos en los que la escala indica la favorabilidad de las condiciones ambientales para una especie y su distribución potencial. Se trata de valores descritos como resultado de los modelos antes mencionados, que resaltan una extensión como zona potencial para las especies.
En México, se ha documentado el impacto del cambio climático en los ecosistemas y las especies forestales. Este puede reducir considerablemente las poblaciones y afecta, en gran medida, su área de distribución (> 50 %; Gutiérrez y Trejo, 2014). En este estudio, se evaluó la distribución potencial en Pinus ayacahuite, P. hartwegii, Abies hickelii, Quercus laurina y Q. ocoteifolia, debido a las modificaciones en temperatura y precipitación, las cuales llegaron al punto en el que se esperan extinciones locales (Guitérrez y Trejo, 2014).
El objetivo de este trabajo es modelar la idoneidad ambiental y la distribución potencial tanto actual como futura de Pseudotsuga menziesii en México, para determinar la influencia del cambio climático en su distribución en ese país.
El área de estudio comprendió la República Mexicana, ubicada entre la región Neártica y Neotropical. México se caracteriza por presentar una amplia variedad de climas a lo largo de su extensión (García, 2004); se conoce que Pseudotsuga menziesii tiene preferencia por zonas templadas del país (Hernández et al., 2005; Ríos et al., 2010), como el Eje Neovolcánico, la Sierra Madre Occidental, la Sierra Madre Oriental y la Sierra Madre del Sur (Figura 1).
Los puntos de presencia de la especie se obtuvieron de literatura especializada (Domínguez et al., 2004; Hernández et al., 2005; Juárez-Agis et al., 2006; López-Upton et al., 2015; Ríos et al., 2010) y de The Global Biodiversity Information Facility (GBIF). Para esta investigación no se consideran plantaciones forestales y solo se tomaron en cuenta registros con una base histórica de 1960-2018. Los datos obtenidos se georreferenciaron en Qgis® para su posterior depuración. Se eliminaron los registros duplicados y espacialmente correlacionados; con tal fin, se utilizó la herramienta NicheToolBox (Osorio-Olvera et al., 2020) y se aplicó una distancia de 3 km entre puntos (Figura 1). Una vez procesada la información, se dividió en 3 conjuntos: 2 para calibrar y crear los modelos candidatos y 1 de datos independientes para evaluar estos modelos (Muscarella et al., 2014).
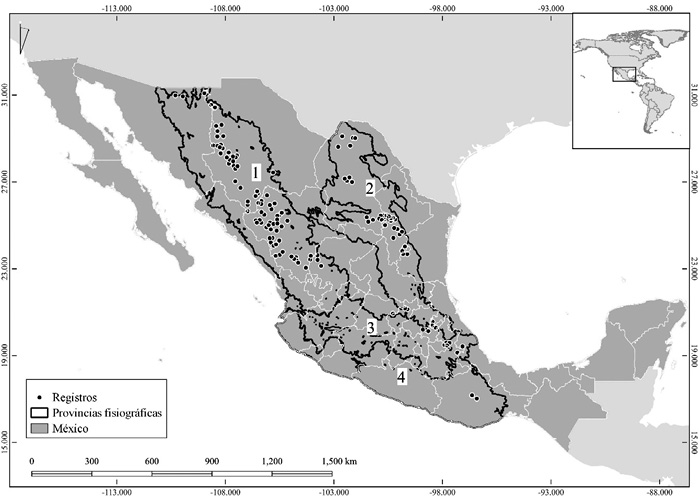
Figura 1. Área de estudio utilizada para la elaboración de modelos de idoneidad ambiental y distribución potencial de Pseudotsuga menziesii a futuro. 1) Sierra Madre Occidental; 2) Sierra Madre Oriental; 3) Eje Neovolcánico; 4) Sierra Madre del Sur.
Figure 1. Study area used for modeling the environmental suitability and potential distribution of Pseudotsuga menziesii in the future. 1) Sierra Madre Occidental; 2) Sierra Madre Oriental; 3) Eje Neovolcánico; 4) Sierra Madre del Sur.
Hecho esto, se trazó un área de calibración (M) basada en la distribución conocida de la especie, en este caso, las zonas accesibles para ella a lo largo de su historia (Cobos et al., 2019). Únicamente, se contempló la distribución en México, debido a que se busca obtener resultados de forma más regional.
Como base para delimitar la M, se emplearon los polígonos de las regiones fisiográficas de México (INEGI 2001). Utilizando solo aquellas con registros de la especie, se consideraron los polígonos de la Sierra Madre Occidental, Sierra Madre Oriental, Sierra Madre del Sur y del Eje Neovolcánico, más un buffer de 10 km como área M, con miras a incluir puntos que se encontraban muy cerca de la línea delimitada por la capa para las provincias.
Para este estudio se utilizaron variables de tipo climático, debido a que se tiene mejor certeza de la tendencia de los cambios en el futuro (Hajima et al., 2020). Las variables ambientales consideradas para la creación del modelo fueron obtenidas de la base de datos WorldClim 2.1, la cual consiste en 19 variables de precipitación y temperatura: BIO1 = temperatura media anual; BIO2 = rango medio diurno (media mensual (temperatura máxima - temperatura mínima)); BIO3 = isotermalidad (BIO2/BIO7) (×100); BIO4 = estacionalidad de la temperatura (desviación estándar × 100); BIO5 = temperatura máxima del mes más cálido; BIO6 = temperatura mínima del mes más frío; BIO7 = rango anual de temperatura (BIO5-BIO6); BIO8 = temperatura media del trimestre más húmedo; BIO9 = temperatura media del trimestre más seco; BIO10 = temperatura media del trimestre más cálido; BIO11 = temperatura media del trimestre más frío; BIO12 = precipitación anual; BIO13 = precipitación del mes más húmedo; BIO14 = precipitación del mes más seco; BIO15 = estacionalidad de las precipitaciones (coeficiente de variación); BIO16 = precipitación del trimestre más húmedo; BIO17 = precipitación del cuarto más seco; BIO18 = precipitación del trimestre más cálido; BIO19 = precipitación del cuarto más frío. Todas ellas son resultado de los valores mensuales de temperatura y precipitación, para generar variables relevantes biológicamente, y representan los patrones, como precipitación anual, estacionalidad y valores extremos de las variables ambientales limitantes. Igualmente, esas variables ambientales contempladas para la elaboración del modelo fueron generadas a partir de datos mensuales históricos para el periodo 1970-2000, con una resolución de 30 segundos (aproximadamente 1 km en el ecuador). En las transferencias a futuro se usó el modelo MIROC-ES2L. Este incorpora un componente biogeoquímico terrestre con interacción explícita carbono-nitrógeno, con el afán de tener en cuenta el control de nutrientes del suelo sobre el crecimiento de las plantas y el sumidero de carbono de la tierra (Hajima et al., 2020). En este estudio, se incluye solo 2 vías socioeconómicas compartidas (SSPs por sus siglas en inglés): 4.5 (escenario de bajas emisiones) y 8.5 (escenario de altas emisiones); estas son proyecciones climáticas futuras reducidas de la Fase del Proyecto de Intercomparación de Modelos Acoplados 6 (CMIP6 por sus siglas en inglés; Hajima et al., 2020). Las transferencias fueron proyectadas hacia 3 periodos (2040, 2060 y 2080), los cuales son valores promediados durante lapsos de 20 años (2020-2040, 2041-2060, 2061-2080); información disponible en http://www.worldclim.org/.
Se efectuó una selección previa de las variables climáticas y se evitó con esto un sobreajuste en los modelos (Raghavan et al., 2019). 1) Se evaluó la trascendencia, utilizando la prueba de Jackknife dentro de MaxEnt (Phillips et al., 2017); 2) se generó una tabla de correlación con ayuda de la herramienta r.covar implementada en Qgis®; 3) se descartaron aquellas variables irrelevantes, con una importancia baja y con una correlación > 0.8 (Muscarella et al., 2014). Como resultado de las 19 variables climáticas para la creación del modelo, se utilizaron 7 (temperatura media; isotermalidad; estacionalidad de la temperatura; temperatura máxima del mes más cálido; precipitación; precipitación del mes más seco; estacionalidad de la precipitación).
Además, se empleó el algoritmo MaxEnt 3.4.1 (Phillips et al., 2017). Para la creación y evaluación de los modelos, se usó el paquete Kuenm, (Cobos et al., 2019) en la plataforma de Rstudio® (https://www.rstudio.com/). Se utilizaron 14 multiplicadores de regularización (regularization multipliers: 0.1, 0.2, 0.3, 0.4, 0.5, 0.6, 0.7, 0.8, 0.9, 1, 2, 3, 4, 5) y la combinación de 15 clases de características entre Linear, Quadratic, Product y Hinge (feature clases: l, q, p, h, lq, lp, lh, qp, qh, ph, lqp, lqh, lph, qph, lqph), lo cual dio como resultado un total de 210 modelos candidatos. Los puntos de presencia se dividieron en subconjuntos para la creación de dichos modelos candidatos; se dejó el 100 % y 70 % en entrenamiento y calibración de modelos candidatos, respectivamente, y el 30 % (presencias independientes) para prueba y selección de los modelos (Cobos et al., 2019).
La evaluación y selección de los modelos candidatos se basó en el rendimiento, la capacidad predictiva y la complejidad (Cobos et al., 2019). Las tasas de significación se calcularon en modelos creados con datos de entrenamiento, con subconjuntos de datos de prueba independientes; esto se valoró mediante el ROC parcial y, seguido, se descartaron todos los modelos para los cuales la tasa de omisión “OR” (por sus siglas en inglés) era ≤ 5 % (Omission Rate; Fielding y Bell, 1997). Por último, se aplicó el criterio de información de Akaike, corregido para tamaños de muestra pequeños (AICc), así como se eligió el modelo con un valor delta AICc ≤ 2 (Cobos et al., 2019; Raghavan et al., 2019). Lo antes mencionado se realizó con ayuda del paquete Kuenm_ceval en R (Cobos et al., 2019).
En este estudio, también se evaluaron dos puntos importantes en la distribución de la especie, la extensión potencial en km2 y la idoneidad ambiental. Se observó la pérdida para cada modelo en el espacio temporal.
1) Se exportó el modelo en un formato de salida tipo logístico, el cual representa valores de idoneidad ambiental (entre 0 y 1) de la distribución de la especie (Jiménez-Salazar y Méndez-González, 2021).
2) Para estimar el área predicha, se generó un modelo de distribución binario. Con tal fin, se utilizó el umbral de sensibilidad y especificidad equivalentes: equal training sensitivity and specificity; posteriormente, se realizó una prueba binomial, usando el umbral previamente evaluado y seleccionado.
Con el propósito de evaluar la influencia del cambio climático en las poblaciones de P. menziesii, se usaron 2 métricas: primero, se valoró la pérdida en km2, con el modelo binario; posteriormente, la disminución de la idoneidad ambiental en porcentaje. Para esto último, se aplicó la siguiente metodología:
Se empleó, como plantilla de corte, los modelos de distribución potencial. Se recortó la idoneidad para cada periodo, con el objetivo de visualizar los cambios de extensión y la pérdida de idoneidad. Consecutivamente, se calculó la disminución de la idoneidad en porcentaje (PPi), mediante la Ecuación 1, de la siguiente manera:
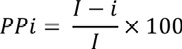 (E. 1)
(E. 1)
• I = idoneidad ambiental en el presente
• i = idoneidad ambiental en el futuro
Este cálculo representa la variación de la idoneidad ambiental en el futuro; los valores positivos indican una disminución de la idoneidad y los negativos, ganancia. La influencia de las variables se interpretó directamente de las curvas de respuesta. Estas muestran cómo aumenta o decrece la idoneidad climática prevista, a medida que cambia cada variable ambiental.
La contribución e importancia de las variables climáticas al modelo se sacó de la tabla que MaxEnt genera a partir del Jackknife. Esto se estima basado en cada iteración del algoritmo con los datos de entrenamiento (Phillips et al., 2017). El valor de importancia se logra permutando aleatoriamente los valores de cada variable climática en los datos de entrenamiento de presencia y fondo. El modelo se vuelve a evaluar con la información permutada y la caída resultante del AUC, este se normaliza en porcentaje (Phillips et al., 2017).
De un total de 1 450 registros disponibles de Pseudotsuga menziesii, solo se utilizaron 138 (Figura 1). El modelo seleccionado se creó a partir de 3 clases de entidad: Linear, Quadratic y Hinge. Se obtuvo un ROC parcial de 1.3, tasa de omisión de 0.09 %, más un delta AICc = 0 y AICc de 2 415.9. El umbral elegido fue 0.3 y la prueba binomial arrojó valores significativos de p-value = 1.734e-11, con un área predicha para el presente de 164 110 km2 (Figura 2; Cuadro 2).
Las variables que mejor predicen la distribución actual de P. menziesii son temperatura media anual (59.5 %) y temperatura máxima del mes más cálido (16.6 %), contribuyendo al modelo con 76.1 %; las que menos aportaron al modelo fueron isotermalidad (0.6 %) y precipitación media anual (0.1 %). De acuerdo con la influencia de las variables, las condiciones óptimas para P. menziesii se ubicarán por debajo de los 16 °C, media anual, por lo que afectan negativamente temperaturas mayores a 30 °C en el mes más cálido (temperatura máxima del mes más cálido), en zonas con una marcada estacionalidad de la temperatura. Aunque las variables de precipitación no presentan una contribución alta (< 7.6), se observó una influencia a regiones de baja precipitación (100-1 500 mm), por lo que se puede inferir que los acumulados anuales de lluvias no representan un factor limitante en la distribución de P. menziesii (Cuadro 1).
Cuadro 1. Contribución, importancia e intervalos óptimos de las variables ambientales en la distribución potencia de Pseudotsuga menziesii, en el presente
Table 1. Contribution, importance, and optimal ranges of environmental variables in the current power distribution of Pseudotsuga menziesii
|
Variable |
Contribución % |
Importancia % |
Intervalos óptimos |
|
Temperatura media anual |
59.5 |
8.5 |
6-16 °C |
|
Temperatura máxima del mes más cálido |
16.6 |
61.5 |
16-30 °C |
|
Estacionalidad de la temperatura |
9.4 |
21.3 |
> 200 |
|
Estacionalidad de la precipitación |
7.6 |
7.4 |
40-100 mm |
|
Precipitación del mes más seco |
6.2 |
0 |
> 50 mm |
|
Isotermalidad |
0.6 |
1.2 |
> 70 |
|
Precipitación media anual |
0.1 |
0.2 |
100-1 500 mm |
Para el escenario actual, se obtuvo lo indicado en el norte del país (Figura 2). Se encontraron 6 zonas de importancia, con alta idoneidad ambiental, que presentan una mayor extensión para la Sierra Madre Occidental, específicamente, en los estados de Chihuahua y Durango, hacia la Sierra Madre Oriental en Nuevo León y Coahuila (Figura 2). Hacia el centro y sur de la República Mexicana, aparecen condiciones idóneas en los estados de Querétaro, Hidalgo y Puebla, dentro del Eje Neovolcánico, mientras que para la Sierra Madre del Sur la idoneidad ambiental disminuyó a valores en el rango 0.3-0.4 y solo hubo condiciones idóneas dentro del estado de Oaxaca, el área más sureña de la distribución potencial y conocida de la especie (Figura 2).
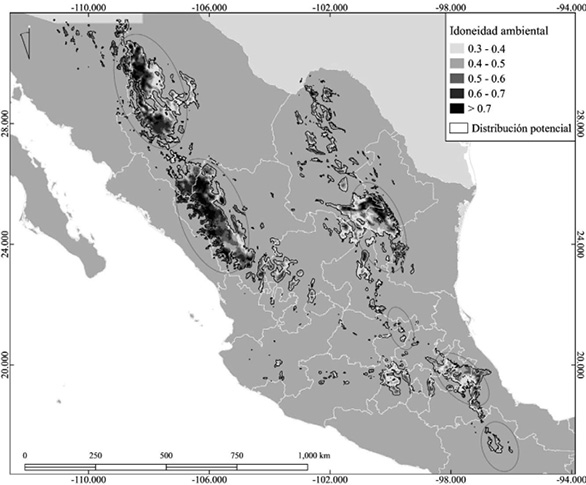
Figura 2. Idoneidad ambiental y distribución potencial de Pseudotsuga menziesii en el presente. Los círculos representan zonas de importancia de acuerdo con lo siguiente: 1) tienen poblaciones de la especie y 2) presentan áreas de alta idoneidad ambiental.
Figure 2. Environmental suitability and potential distribution of Pseudotsuga menziesii at present. The circles represent the areas of importance according to: 1) having populations of the species, and 2) presenting areas of high environmental suitability.
3.1 Impacto del cambio climático
Pseudotsuga menziesii presentó una mayor preferencia por zonas templadas del país (2040; 2060; 2080), acorde con la respuesta de las variables y bajo un escenario de cambio climático. Se infiere que el aumento de la temperatura media anual sobre la superficie terrestre será el factor climático que más afecte la idoneidad ambiental y distribución de la especie (Cuadro 1).
Según los escenarios de cambio climático, en el 2040, el área de distribución potencial se reduce en el norte de México; los estados de Chihuahua, Coahuila y Nuevo León serían los más afectados, con una pérdida total de 50.7 % a 49.6 %, para los SSPs 4.5 y 8.5, respectivamente (Figura 3; Cuadro 2).
En el 2060, la distribución potencial se reducirá 63.4 % (SSPs 4.5) y 71.9 % (SSPs 8.5), afectará el norte y parte del centro del país, perderá extensión en los estados de Querétaro, Hidalgo y Puebla (Figura 3; Cuadro 2).
Por último, para el 2080, se observa un escenario grave, habrá una pérdida de 74.4 % (SSPs 4.5) y 92.6 % (SSPs 8.5); las poblaciones de Chihuahua, Coahuila, Nuevo León y Oaxaca serían las más afectadas en el SSPs 8.5 (Figura 3; Cuadro 2).
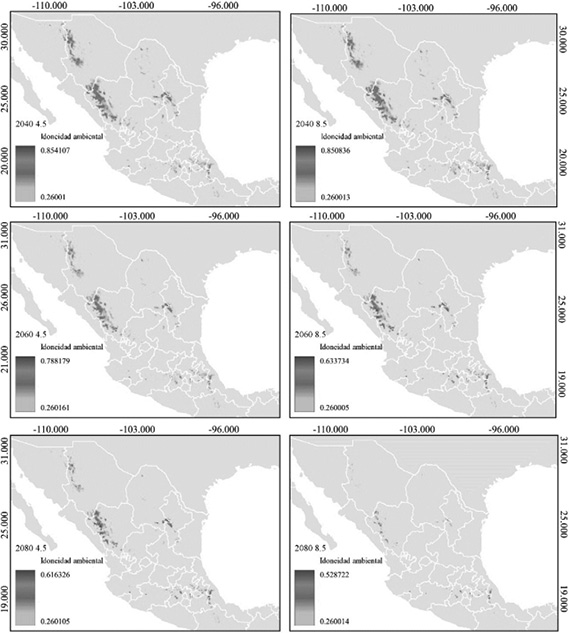
Figura 3. Idoneidad ambiental de Pseudotsuga menziesii en 3 contextos y bajo 2 escenarios, 4.5 (escenarios de bajas emisiones) y 8.5 (escenario de altas emisiones)
Figure 3. Environmental suitability of Pseudotsuga menziesii in 3 context and under 2 scenarios, 4.5 (low emissions scenarios) and 8.5 (high emissions scenario)
Cuadro 2. Efecto del cambio climático en la distribución de Pseudotsuga menziesii. Se presentan las áreas estimadas, a partir de modelo binario en km2 y su porcentaje de pérdida respecto al presente.
Table 2. Effect of climate change on the distribution of Pseudotsuga menziesii. The estimated areas from the binary model are presented in km2 and their percentage of loss with respect to the present.
|
Escenario |
Presente |
2040 |
2060 |
2080 |
|||
|
Ambiente |
4.5 |
8.5 |
4.5 |
8.5 |
4.5 |
8.5 |
|
|
Superficie (km2) |
164 110 |
80 909.3 |
82 626.9 |
60 006.9 |
46 121.3 |
41 908.2 |
12 101.7 |
|
Porcentaje de pérdida (%) |
50.7 |
49.6 |
63.4 |
71.9 |
74.4 |
92.6 |
|
En cuanto a los cambios en la idoneidad ambiental de la especie, para el 2040, de acuerdo con el modelo SSPs 4.5, las zonas que más presentan esta característica (I = > 0.6) disminuyeron hasta el 50 % en el norte (Chihuahua, Coahuila y Nuevo León). En contraste, en el centro y sur del país, las modificaciones no son tan evidentes, con una reducción de la idoneidad entre 25 y 2 %; para el SSPs 8.5, se espera una disminución máxima de 52 % y una mínima de 0.1 %.
Proyectando el 2060, la idoneidad ambiental se redujo 59 % (SSPs 4.5) y 64 % (SSPs 8.5) en sus valores máximos, con afectaciones significativas en los estados del norte del país. Destaca que las zonas donde no se observa un porcentaje de variación considerado (1 y 25 %) son aquellas localizadas en las áreas altas de la Sierra Madre Occidental, Sierra Madre Oriental, así como en el centro y sur de la nación, específicamente, en los estados de Puebla, Hidalgo, Tlaxcala y parte de Oaxaca, para los SSPs 4.5 y SSPs 8.5 (Figura 3; Figura 4).
Finalmente, para el 2080, se observa una reducción de la idoneidad ambiental de 64 % en el SSPs 4.5 y de 68 % en el SSPs 8.5, último escenario donde la constante es la disminución tanto en el sur como en el norte del país (Figura 4).
Se apreció que las regiones las cuales se mantienen potencialmente tolerantes al cambio climático se ubican en las zonas altas de la Sierra Madre Occidental, Sierra Madre Oriental y del Eje Neovolcánico (Figura 3), ya que el porcentaje de variación en ellas osciló entre los siguientes rangos, respectivamente: 0.1-22 % (2040), 2-25 % (2060) y -0.5-25 % (2080).
Se observó, con respecto a la distribución potencial y la reducción de la idoneidad ambiental, que conforme aumenta el periodo en las zonas donde se presentaban condiciones óptimas ellas se ven más afectadas que las de baja idoneidad (I = 0.3-0.4; Figura 2). Notamos que durante los periodos de 2040 y 2060 los cambios en el centro y sur de la distribución no aumentaron más del 27 % (Figura 4).
Las coníferas se han visto favorecidas por su tolerancia climática al frío extremo y veranos calurosos. Reich y Oleksyn (2008) mencionan que incluso un calentamiento climático moderado influirá en la supervivencia y el crecimiento; por ejemplo, las poblaciones del centro y sur muestran una supervivencia de hasta 80 % a -4 ºC y esta disminuyó, en algunos casos, hasta 20 % a temperaturas de 6 ºC. Por ello, es importante mantener una constante actualización de las bases de datos del clima y, a su vez, de las afectaciones que este trae a las especies forestales.
En general, se muestra un panorama desalentador para Pseudotsuga menziesii; en las próximas décadas, la especie podría presentar un alto riesgo de extinción de forma local. Los resultados indican la pérdida de idoneidad ambiental para el norte, la cual descenderá en un alto porcentaje (Figura 4); esto fuertemente influenciado por las condiciones climáticas que predominan, en particular templadas, lo que produce un efecto de aislamiento, reducción y de desplazamiento hacia zonas de mayor elevación (Ledig et al., 2010).
Resultados obtenidos por Bansal et al. (2015) indican que el aumento de las temperaturas otoñales reduce la resistencia al frío, lo cual presenta un mayor riesgo de mortalidad en las plántulas. Por ello, el cambio drástico en las temperaturas más norteñas afectaría severamente la especie. Este mismo efecto se observó con Pinus sylvestris en Europa; por debajo de la latitud 62° N, la supervivencia decreció y disminuyó el crecimiento, por debajo de 54° N de latitud (Reich y Oleksyn, 2008).
En la actualidad, estamos viviendo grandes cambios potencializados, principalmente por factores antrópicos, gran parte de las especies forestales en México se verán afectadas por las modificaciones climáticas. Guitérrez y Trejo (2014) mencionan que especies como Pinus hartwegii, Abies hickelii y Quercus ocoteifolia se verán gravemente impactadas, con un decrecimiento en su distribución de 80, 79 y 69 %, respectivamente, para el 2050. Estos resultados se asemejan a nuestras proyecciones y debido a que esas especies cohabitan en parte de su distribución, es posible que pudieran llegar a sufrir una pérdida similar a la de P. menziesii, para el 2080.
De acuerdo con la respuesta de la variable de mayor importancia (temperatura media anual y temperatura máxima del mes más cálido), P. menziesii muestra una preferencia por climas templados con temperaturas entre 6 y 16 °C. Esto coincide con los resultados de Ríos et al. (2010), quien señala que P. menziesii habita en localidades donde la temperatura media oscila entre 10.1 y 14.0 °C. Aunado a esto, lo indagado por Bansal et al. (2015) proporciona bases sólidas para decir que el aumento en la temperatura invernal afectará, en gran medida, las poblaciones de la especie de estudio.
Las variables de precipitación representan una baja importancia para la especie investigada (Cuadro 1). Esto es más evidente en poblaciones de P. menziesii donde existen inviernos relativamente fríos y veranos áridos, ya que aquellas tienen un mayor grado de adaptación a condiciones muy secas, lo cual favorece a los grupos poblacionales futuros ante las sequías de algunas zonas del país (Sheel Bansal et al., 2015).
De acuerdo con los resultados de este trabajo, la precipitación no es una variable que influya, en gran manera, en la distribución de P. menziesii. Ríos, (2010) aluden a que los acumulados anuales de precipitaciones no representan un factor limitante para el desarrollo de pinares. Sin embargo, con periodos extensos de sequía, las poblaciones pueden caer en estrés hídrico, debido a que los pinos no poseen reposo vegetativo, así que una larga y extensa sequía podría comprometer su existencia (Martínez-Vilalta et al., 2012).
El cambio de temperatura en los diferentes periodos indica una tendencia de aumento, que desfavorece mucho las poblaciones de P. menziesii, principalmente del norte (Figura 4). Lo antes especificado se debe a que el crecimiento del ancho de los anillos se beneficia con la presencia de bajas temperaturas (Arreola-Ortiz et al., 2010).
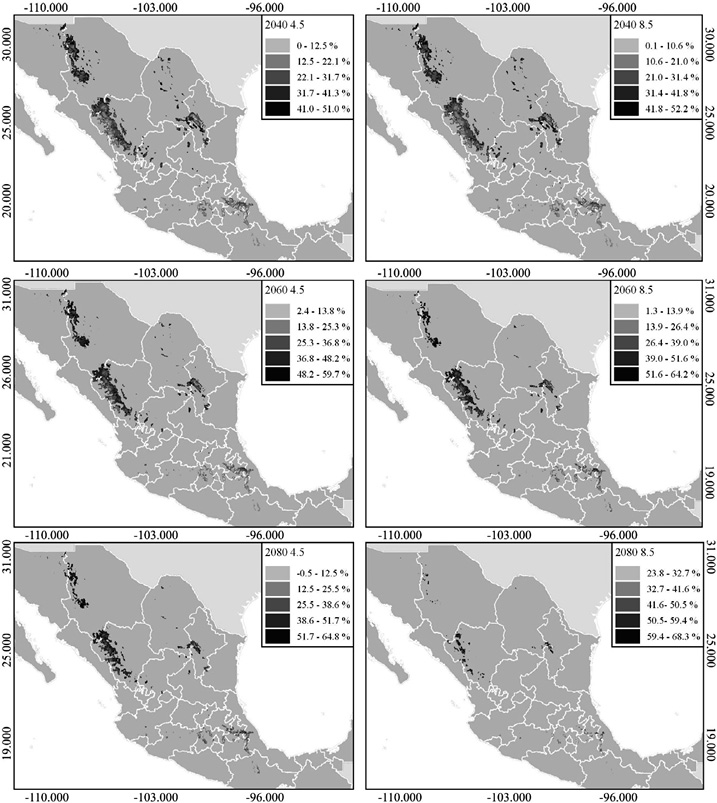
Figura 4. Porcentaje de pérdida de idoneidad ambiental en 3 futuros y bajo 2 escenarios, 4.5 (escenarios de bajas emisiones) y 8.5 (escenario de altas emisiones)
Figure 4. Percentage of loss of environmental suitability in 3 futures and under 2 scenarios, 4.5 (low emissions scenarios) and 8.5 (high emissions scenario)
El cambio climático afecta en diferente grado las distintas poblaciones de P. menziesii (Hernández et al., 2005). Observamos, hacia el 2040 y 2060, que la idoneidad disminuye más del 50 % para las poblaciones norteñas (Figura 4), las primeras en verse afectadas por el cambio climático y, aunque aquella en el presente es baja para las del sur, estas muestran una adaptación local que les permite cierta resistencia ante estos cambios hacia temperaturas más cálidas (Kremer et al., 2014). Por ello, es posible que dichas poblaciones del sur tengan mayor probabilidad y viabilidad para transferir semillas, cuando las condiciones climáticas del norte sean desfavorables (Hernández et al., 2005); con esto, el movimiento de la semilla de manera altitudinal, en un sentido positivo, podría ayudar a disminuir y prevenir efectos negativos, frente una baja adaptación, en un futuro escenario de calentamiento global (Sáenz-Romero et al., 2004).
La distancia entre las diferentes poblaciones genera un mayor riesgo de extinción; las aisladas tienen un menor flujo genético, lo cual puede producir una variabilidad genética reducida (Juárez-Agis et al., 2006; Ledig et al., 2010). Aunado a esto, la baja densidad aumenta ese riesgo de extinción, ya que una población pequeña igual tiene bajo flujo genético y está sujeta a una deriva genética aleatoria. Con base en lo anterior, grupos poblacionales marginados del centro y sur de la distribución natural de P. menziesii son más propensos a desaparecer por su aislamiento (López-Upton et al., 2015). Sin embargo, estas agrupaciones presentan mayor resistencia a los escenarios de cambio, por lo que se necesita diseñar estrategias que permitan el intercambio genético entre poblaciones. Así, al considerar la migración de semilla o movimientos de germoplasma, se debe efectuar estudios que establezcan los rangos de movimiento (Castellanos-Acuña et al., 2014).
Los resultados de este trabajo indican que las poblaciones más sureñas no se verán afectadas, en gran medida, con reducciones en la idoneidad ambiental, de 0.1-22 % (2040), 2-25 % (2060) y 0.5-25 % (2080). Estas podrían presentar mayor resistencia a los escenarios de cambio; sin embargo, para tal aseveración, se deben hacer análisis más locales, con el objetivo de diseñar estrategias que permitan el intercambio genético entre poblaciones, contemplando lo reportado por autores como Ríos et al. (2010), quienes, antes de transferir semillas o plantas entre zonas con condiciones ambientales distintas, hacen pruebas o ensayos de procedencia. No obstante, ante un escenario de cambio climático y una baja adaptación, la transferencia de semillas pudiera ser una solución, siempre y cuando las condiciones de los sitios tengan cierto grado de similitud o cercanía (Bansal et al., 2015; Ríos et al., 2010; Sáenz-Romero et al., 2004).
Las variables climáticas que predijeron la distribución de P. menziesii son la temperatura media anual (59.5 %) y la temperatura máxima del mes más cálido (16.6 %), contribuyendo al modelo con 76.1 %; siendo estas variables de relevancia ante el escenario de cambio climático.
Algo que se observa en los modelos de este trabajo es que, a pesar de que actualmente el norte de México mantiene una alta idoneidad ambiental (I = > 0.5), a futuro, estas zonas serán las más afectadas con una disminución mayor al 50 %.
En los diferentes escenarios, se nota una pérdida drástica de superficie, con lo que la prevalencia de la especie pudiera encontrarse en detrimento. La pérdida de zonas de idoneidad ambiental es evidente, esto se observa en las superficies derivadas de los modelos. Ante el escenario de calentamiento global, las poblaciones más susceptibles se encuentran en los estados del norte y, a pesar de que las del sur de México (Oaxaca) presentan un mejor resultado para los escenarios del 2040 y 2060, estas tienden a la disminución o desaparición en el escenario de las de SSPs 8.5, en el 2080. Los posibles escenarios ayudan establecer estrategias para la preservacion de la especie en zonas de su distribucion actual
Las personas autoras declaran que han cumplido con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en la producción del manuscrito; que no hay conflictos de intereses de ningún tipo; que todas las fuentes financieras se mencionan completa y claramente en la sección de agradecimientos; y que están totalmente de acuerdo con la versión final editada del artículo.
Se agradece a la revista y las personas revisoras anónimas, por sus valiosos aportes al documento.
Álvarez-Moctezuma, J. G., Alia-Tejacal, I., Colinas-León, M. T. y Sahagún-Castellanos, J. (2007). Interspecific differences in postharvest quality on Mexican Christmas trees. Silvae Genetica, 56(1-6), 65-73. https://doi.org/10.1515/sg-2007-0010
Arreola-Ortiz, M. R. y Návar-Cháidez, J. D. J. (2010). Análisis de sequías y productividad con cronologías de Pseudotsuga menziesii Rob. & Fern., y su asociación con El Niño en el nordeste de México. Investigaciones geográficas, (71), 7-20.
Bansal, S., St. Clair, J. B., Harrington, C. A. y Gould, P. J. (2015). Impact of climate change on cold hardiness of Douglas-fir (Pseudotsuga menziesii): environmental and genetic considerations. Global Change Biology, 21(10), 3814-3826. https://doi.org/10.1111/gcb.12958
Castellanos-Acuña, D., Lindig-Cisneros, R. A., Silva-Farias, M. Á. y Sáenz-Romero, C. (2014). Zonificación altitudinal provisional de Abies religiosa en un área cercana a la reserva de la biósfera de la Mariposa Monarca, Michoacán. Revista Chapingo serie ciencias forestales y del ambiente, 20(2), 215-225. https://doi.org/10.5154/r.rchscfa.2013.11.041
Cobos, M. E., Peterson, A. T., Barve, N. y Osorio-Olvera, L. (2019). Kuenm: an R package for detailed development of ecological niche models using Maxent. PeerJ, 7, e6281. https://doi.org/10.7717/peerj.6281
DOF. (2019). Modificación del Anexo Normativo III, Lista de especies en riesgo de la Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección ambiental - Especies nativas de México de flora y fauna silvestres - Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio - Lista de especies en riesgo, publicada el 30 de diciembre de 2010. Consultado en línea: www.dof.gob.mx/nota_detalle.php?codigo=5578808&fecha=14/11/2019#gsc.tab=0.
Domínguez A., F. A., Vargas H., J. J., López U., J., Ramírez V., P. y Guízar N., E. (2004). Caracterización ecológica de Pseudotsuga menziesii en Pinal de Amoles, Querétaro: nueva población natural en México. Anales del Instituto de Biología, 75(2), 191-203.
Escoto Castillo, A., Sánchez Peña, L. y Gachuz Delgado, S. (2017). Trayectorias Socioeconómicas Compartidas (SSP): nuevas maneras de comprender el cambio climático y social. Estudios demográficos y urbanos, 332(3), 669-693. https://doi.org/10.24201/edu.v32i3.1684
Fielding, A. H. y Bell, J. F. (1997). A review of methods for the assessment of prediction errors in conservation presence/absence models. Environmental Conservation, 2, 38-49. https://doi.org/10.1017/S0376892997000088
García, E. (2004). Modificaciones al Sistema de Clasificación Climática de Köppen. Instituto de Geografía. Universidad Nacional Autónoma de México. Scientia Horticulturae, 140, 131-139.
García, S. A. G., Rodríguez, E. A., Calderón, O. A., Garza, E. T. y Ávila, G. G. (2021). Contenido de carbono y estructura horizontal de un bosque templado en Guadalupe y Calvo, Chihuahua. Revista Mexicana de Ciencias Forestales, 12(63). https://doi.org/10.29298/rmcf.v12i63.800
Guitérrez, E. y Trejo, I. (2014). Efecto del cambio climático en la distribución potencial de cinco especies arbóreas de bosque templado en México. Revista Mexicana de Biodiversidad, 85(1), 179-188. https://doi.org/10.7550/rmb.37737
Hajima, T., Watanabe, M., Yamamoto, A., Tatebe, H., Noguchi, M. A., Abe, M. y Kawamiya, M. (2020). Development of the MIROC-ES2L Earth system model and the evaluation of biogeochemical processes and feedbacks. Geoscientific Model Development, 13, 2197-2244. https://doi.org/10.5194/gmd-13-2197-2020
Hernández, V. J. R., Hernández, J. J., Upton, J. L. y Huerta, H. V. (2005). Variación morfológica y anatómica en poblaciones mexicanas de Pseudotsuga (Pinaceae). Acta Botánica Mexicana, (70), 47-67. https://doi.org/10.21829/abm70.2005.987
INEGI. (2001). Conjunto de Datos Vectoriales Fisiográficos. Continuo Nacional. Escala 1:1’000,000. Serie I. Disponible en línea: https://www.inegi.org.mx/app/biblioteca/ficha.html?upc=702825267575
Jiménez-Salazar, M. Á. y Méndez-González, J. (2021). Actual and potential distribution of Pinus engelmannii Carriére under climate change scenarios. Madera y Bosques, 27(3). https://doi.org/10.21829/myb.2021.2732117
Juárez-Agis, A., López-Upton, J., Vargas-Hernández, J. J. y Sáenz-Romero, C. (2006). Variación geográfica en la germinación y crecimiento inicial de plántulas de Pseudotsuga menziesii de México. Agrociencia, 40(6), 783-792.
Kremer, A., Potts, B. M. y Delzon, S. (2014). Genetic divergence in forest trees: understanding the consequences of climate change. Functional Ecology, 28(1), 22-36. https://doi.org/10.1111/1365-2435.12169
Ledig, F. T., Rehfeldt, G. E., Sáenz-Romero, C. y Flores-López, C. (2010). Projections of suitable habitat for rare species under global warming scenarios. American Journal of Botany, 97(6), 970-987. https://doi.org/10.3732/ajb.0900329
López-Upton, J., Valdez-Lazalde, J. R., Ventura-Ríos, A., Vargas-Hernández, J. J. y Guerra-de-la-Cruz, V. (2015). Extinction risk of Pseudotsuga menziesii populations in the central region of Mexico: An AHP analysis. Forests, 6(5), 1598-1612. https://doi.org/10.3390/f6051598
Martínez-Vilalta, J., Auadé, D., Banqué, M., Barba, J., Curiel Yuste, J., Galiano, L., García, N., Gómez, M., Hereş, A., López, B., Lloret, F., Poyatos, R., Retana, J., Sus, O., Vayreda, J. y Vilà-Cabrera, A. (2012). Las poblaciones ibéricas de pino albar ante el cambio climático: con la muerte en los talones. Ecosistemas, 21(3), 15-21. https://doi.org/10.7818/ECOS.2012.21-3.03
Monterroso-Rivas, A. I., Gómez-Díaz, J. D. y Tinoco-Rueda, J. A. (2013). Bosque mesófilo de montaña y escenarios de cambio climático: una evaluación en Hidalgo, México. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 19(1), 29-43. https://doi.org/10.5154/r.rchscfa.2012.03.029
Muscarella, R., Galante, P. J., Soley‐Guardia, M., Boria, R. A., Kass, J. M., Uriarte, M. y Anderson, R. P. (2014). ENM eval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods in Ecology and Evolution, 5(11), 1198-1205. https://doi.org/10.1111/2041-210X.12261
Osorio-Olvera, L. et al. (2020). ntbox: an R package with graphical user interface for modeling and evaluating multidimensional ecological niches. Methods in Ecology and Evolution, 11(10), 1199-1206. https://doi.org/10.1111/2041-210X.13452
Peterson, T., Sánchez-Cordero, V., Soberón, J., Bartley, J., Buddemeier, R. y Sánchez-Navarro, A. (2001). Effects of global climate change on geographic distributions of Mexican Cracidae. Ecological Modeling, 144(1), 21-30. https://doi.org/10.1016/S0304-3800(01)00345-3
Plasencia-Vázquez, A. H., Vílchez-Ponce, C. R., Ferrer-Sánchez, Y. y Veloz-Portillo, C. E. (2022). Efecto del cambio climático sobre la distribución potencial del hongo Moniliophthora roreri y el cultivo de cacao (Theobroma cacao) en Ecuador continental. Terra Latinoamericana, 40. https://doi.org/10.28940/terra.v40i0.1151
Phillips, S. J., Anderson, R. P., Dudík, M., Schapire, R. E. y Blair, M. E. (2017). Opening the black box: An open‐source release of Maxent. Ecography, 40, 887-893. https://doi.org/10.1111/ecog.03049
Raghavan, R. K., Barker, S. C., Cobos, M. E., Barker, D., Teo, E. J. M., Foley, D. H. y Peterson, A. T. (2019). Potential spatial distribution of the newly introduced long-horned tick, Haemaphysalis longicornis in North America. Scientific reports, 9(1), 498. https://doi.org/10.1038/s41598-018-37205-2
Reich, P. B. y Oleksyn, J. (2008). Climate warming will reduce growth and survival of Scots pine except in the far north. Ecology letters, 11(6), 588-597. https://doi.org/10.1111/j.1461-0248.2008.01172.x
Reynoso Santos, R., Pérez Hernández, M. J., López Báez, W., Hernández Ramos, J., Muñoz Flores, H. J., Cob Uicab, J. V., y Reynoso Santos, M. D. (2018). El nicho ecológico como herramienta para predecir áreas potenciales de dos especies de pino. Revista Mexicana de Ciencias Forestales, 9(48), 47-68. https://doi.org/10.29298/rmcf.v8i48.114
Ríos, A. V., Upton, J. L., Hernández, J. J. V. y De la Cruz, V. G. (2010). Caracterización de Pseudotsuga menziesii (MIRB.) Franco en el centro de México. Implicaciones para su conservación. Revista Fitotecnia Mexicana, 33(2), 1107-116. https://doi.org/10.35196/rfm.2010.2.107
Romero-Sánchez, M. E. (2016). Escenarios de Cambio Climático en el sector forestal. Revista Mexicana de Ciencias Forestales, 7(37), 4-6. https://doi.org/10.29298/rmcf.v7i37.47
Sáenz-Romero, C., Vargas-Hernández, J. J., Bermejo-Velázquez, B. y Ledig, F. T. (2004). Zonificación estatal y altitudinal para la colecta y movimiento de semillas de coníferas en México. Manejo de recursos genéticos forestales, 72-86.
Sheel, B., Harrington, C. A., Gould, P. J. y St. Clair, J. B. (2015). Climate‐related genetic variation in drought‐resistance of Douglas‐fir (Pseudotsuga menziesii). Global Change Biology, 21(2), 947-958. https://doi.org/10.1111/gcb.12719
1 Estudiante, Maestría en Ciencias en Recursos Naturales y Desarrollo Rural, El Colegio de la Frontera Sur (ECOSUR), San Cristóbal de Las Casas, México. rodolfo.rodriguez@posgrado.ecosur.mx; https://orcid.org/0000-0002-3654-1429
2 Docente investigador, Escuela Superior de Ciencias Ambientales, Universidad Autónoma de Guerrero, Acapulco, México. 13457@uagro.mx; https://orcid.org/0000-0001-8839-112
3 Docente investigador, Escuela Superior de Ciencias Ambientales, Universidad Autónoma de Guerrero; Acapulco, México. 13655@uagro.mx; https://orcid.org/0000-0002-3641-3267
4 Docente investigador, Escuela Superior de Ciencias Ambientales, Universidad Autónoma de Guerrero; Acapulco, México. 11266@uagro.mx; https://orcid.org/0000-0003-0021-1767
5 Técnico académico, Instituto de Biología, Universidad Nacional Autónoma de México, Ciudad de México, México. eortiz@ib.unam.mx; https://orcid.org/0000-0003-2932-5098
6 Pasante de Biología, Instituto de Biología, Universidad Nacional Autónoma de México, Ciudad de México, México. rociorrb13@ciencias.unam.mx; https://orcid.org/0000-0001-8141-0586



Escuela de Ciencias Ambientales,
Universidad Nacional, Campus Omar Dengo
Apartado postal: 86-3000. Heredia, Costa Rica
Teléfono: +506 2277-3688
Correo electrónico revista.ambientales@una.ac.cr