
e-ISSN: 2215-3896.
(Julio-Diciembre, 2024). Vol 58(2)
DOI: https://doi.org/10.15359/rca.58-2.1
Open Acces: www.revistas.una.ac.cr/ambientales
e-Mail: revista.ambientales@una.cr
Licencia: CC BY NC SA 4.0
Caracterización acústica de fincas bajo pago por servicios ambientales en la Reserva Forestal Golfo Dulce, Costa Rica
Acoustic characterization of private lands under payment for environmental services at Golfo Dulce Forest Reserve, Costa Rica
Jimmy Barrantes-Madrigal1, Mónica Retamosa-Izaguirre2, Ricardo Bedoya-Arrieta3, Adriana Valerio-Madrigal4, Jonathan Sánchez-Alpízar5
[Recibido: 30 de agosto del 2023, Aceptado: 14 de noviembre del 2023, Corregido: 12 de enero del 2024,
Publicado: 5 de febrero del 2024]
Resumen
[Introducción]: En Latinoamérica, la sensible biodiversidad tropical que se encuentra fuera de las áreas silvestres protegidas está bajo amenaza. En Costa Rica, el pago por servicios ambientales (PSA) representa un mecanismo para proteger bosques en fincas privadas, pero carece de indicadores específicos sobre el estado de la biodiversidad y de conservación de los bosques. [Objetivo]: Analizar el uso del monitoreo acústico para la evaluación de ecosistemas en 11 sitios en la Península de Osa: 9 en fincas privadas y 2 sitios dentro del Parque Nacional Corcovado. [Metodología]: Se grabaron paisajes sonoros del 25 de mayo al 01 de junio del 2021, utilizando grabadoras acústicas. Se estudia el comportamiento de 6 índices acústicos en cada sitio de muestreo y se aplicó un análisis de detección automatizada de vocalizaciones de 2 especies de aves representativas de la zona. [Resultados]: Se encuentra que los sitios más fragmentados presentaron mayor actividad, energía y diversidad acústica, posiblemente por efectos de borde y ecotonos presentes, así como un alto componente de bosque en la matriz circundante. La detección automatizada de vocalizaciones indicó que la especie Hylopezus perspicillatus resultó ser más sensible a las diferencias en el paisaje que Trogon bairdii. [Conclusiones]: El uso de índices acústicos, aunque asocia rasgos del grado de conservación del bosque, no resultó un elemento diferenciador de la calidad del ecosistema en las fincas evaluadas, pero el monitoreo de especies mediante la detección de sus vocalizaciones sí resultó promisorio. Es recomendable un diseño de muestreo metódico y riguroso, con más puntos de muestreo por sitio, para lograr mediciones precisas y evitar sesgos en el análisis.
Palabras clave: conservación; detección automatizada; índices acústicos; monitoreo acústico; paisaje sonoro.
Abstract
[Introduction]: In Latin America, the sensitive tropical biodiversity found outside of protected wilderness areas is under threat. In Costa Rica, the payment for environmental services (PSA) represents a mechanism to protect forests on private farms, but it lacks specific indicators on the state of biodiversity and forest conservation. [Objective]: To analyze the use of acoustic monitoring for ecosystem assessment at 11 sites in the Osa Peninsula, nine on private farms, and two sites within Corcovado National Park. [Methods]: Soundscapes were recorded from May 25 to June 1, 2021, using acoustic recorders. It was analyzed the behavior of six acoustic indices at each sampling site and applied an automated detection analysis of vocalizations of two representative bird species of the area. [Results]: It was found that the most fragmented sites presented higher activity, energy, and acoustic diversity, possibly due to edge effects and ecotones present, as well as a high forest component in the surrounding matrix. Automated vocalization detection indicated that the species Hylopezus perspicillatus turned out to be more sensitive to differences in the landscape than Trogon bairdii. [Conclusions]: The use of acoustic indices, although it associates features of the degree of conservation, was not a differentiating element of the quality of the ecosystem in the evaluated farms. The monitoring of species through the detection of their vocalizations was promising. A careful and rigorous sampling design, with more sampling points per site, is recommended to achieve accurate measurements and avoid bias in the analysis.
Keywords: acoustic indices; acoustic monitoring; automatic detection; conservation; soundscape.
Costa Rica cuenta con uno de los sistemas de áreas protegidas más exitosos de Latinoamérica (Elbers, 2011, González-Maya et al., 2015). No obstante, un gran porcentaje de la biodiversidad se encuentra fuera de dicho sistema, en remanentes boscosos de fincas privadas que funcionan como puente y hábitat para muchas especies (Pringle, 2017).
Ante esto, se han desarrollado iniciativas que fomentan la conservación de bosques en tierras privadas para evitar el cambio de uso de suelo y reducir las tasas de deforestación, como es el caso del pago por servicios ambientales (Porras y Chacón-Cascante, 2018). Por ejemplo, en Costa Rica, el Fondo Nacional de Financiamiento Forestal (FONAFIFO) ha desarrollado un programa por más de 25 años de forma exitosa y con reconocimiento internacional (Sánchez-Chaves y Navarrete-Chacón, 2017). Sin embargo, cuenta con un enfoque general y requiere nueva orientación para identificar servicios ecosistémicos específicos, como el de biodiversidad.
Así, nace en FONAFIFO un proyecto piloto en la Península de Osa, denominado Bosque Vivo y basado en la presencia de fauna terrestre, con especial atención en especies amenazadas y en peligro de extinción, así como aquellos considerados elementos focales de manejo según el Sistema Nacional de Áreas de Conservación (SINAC). Sin embargo, aunque en Costa Rica se han realizado esfuerzos importantes para identificar y catalogar especies (Bermúdez-Rojas y Obando-Acuña, 2021), aún existen muchos vacíos de información sobre la efectiva presencia, distribución y patrones de movimiento de especies en los diferentes sitios y ecosistemas. Para el caso de los esquemas de PSA, esto dificulta la efectiva identificación de zonas prioritarias para la conservación.
Ante esto, se deben valorar nuevos esquemas de evaluación y medición, que generen registros e indicadores de biodiversidad o referentes al estado de conservación de los ecosistemas. El uso de sistemas automatizados puede representar una vía efectiva que facilite no solo la identificación de áreas, sino el seguimiento temporal de los servicios ecosistémicos, lo cual representa una posible herramienta que oriente acciones de conservación.
Una de estas herramientas es el monitoreo acústico-pasivo, una herramienta costo/efectiva para el monitoreo de especies y ecosistemas. Esta técnica se basa en analizar los sonidos del bosque para extraer información sobre el paisaje acústico y las especies sonoras presentes. Los datos obtenidos mediante el monitoreo han sido satisfactoriamente implementados en la evaluación de biodiversidad y en el estado de conservación de ecosistemas (Bradfer‐Lawrence et al., 2019), así como en la obtención de indicadores como abundancia, presencia y comportamiento de especies sonoras en días, semanas o meses enteros de muestreo (Pérez-Granados y Traba, 2021).
En los países de América Latina y el mundo, donde se efectúa el pago por servicios ambientales, la protección de la biodiversidad generalmente es uno de los objetivos de la implementación de este sistema. Sin embargo, una de las problemáticas de incluir este elemento es la dificultad para definir qué es biodiversidad en un contexto de evaluación de servicios ecosistémicos (Ezzine de Blas et al., 2011). Al tener esto en cuenta, el enfoque de monitoreo acústico es un enfoque promisorio como proxi para evaluar la biodiversidad, considerando diferentes aspectos de esta, así como también brindar la posibilidad de enfocarse en especies clave de manera simultánea (las cuales puedan ser identificadas mediante sonidos).
El objetivo de este trabajo fue evaluar la posibilidad de establecer la calidad del ecosistema boscoso existente en las fincas privadas sometidas a pago por servicios ambientales en la Reserva Forestal Golfo Dulce, mediante una caracterización y análisis del paisaje sonoro. Se realizó una caracterización de los sitios de estudios de acuerdo con su estructura vegetal y variables de paisaje en un área circundante y se comparó los valores de índices acústicos entre sitios de estudio con el objetivo de identificar patrones y su relación con las características de cada lugar. Además, se realizó un análisis de detección automatizada de dos especies presentes en el área de estudio. Finalmente se brindan recomendaciones sobre el uso de los índices acústicos en la evaluación de las fincas bajo pago por servicios ambientales. Se espera que este trabajo contribuya a proveer herramientas y recomendaciones para la evaluación de fincas privadas mediante métodos acústicos de manera costo-efectiva, que sean de provecho para la toma de decisiones a escala local y nacional.
Se seleccionaron 11 sitios con remanentes de bosque maduro. De estos, 9 corresponden a fincas privadas bajo pago por servicios ambientales en la Reserva Forestal Golfo Dulce y 2 sitios dentro del Parque Nacional Corcovado (Cuadro 1; Figura 1).
Cuadro 1. Descripción de los sitios de muestreo seleccionados para el análisis del paisaje sonoro.
Table 1. Description of the selected sampling sites for the soundscape analysis.
|
Código |
Condición |
Detalle |
|
CRRI |
Finca privada (PSA) |
Parte alta de una loma, elevación de 88 m s. n. m., dominada por una cobertura de bosque maduro con presencia de árboles de gran tamaño, pero que no cierran en su totalidad el dosel. Sotobosque denso con muchas palmas. Topografía ondulada. Afluente de agua aproximadamente a 100 m. Poca influencia antrópica. |
|
HEMO |
Finca privada (PSA) |
Parte media de una loma, elevación de 170 m s. n. m., con cobertura de un bosque maduro con presencia de árboles de gran tamaño, pero con presencia de espacios. Sotobosque abierto. Topografía ondulada. Afluente de agua más cercano a 250 m aproximadamente. Influencia antrópica media. |
|
JUCU |
Finca privada (PSA) |
Pequeño valle en la parte media de una loma, elevación de 334 m s. n. m., con cobertura de un bosque maduro con presencia de árboles de gran tamaño y palmas de porte medio, pero con algunos claros de bosque. Sotobosque de intermedia densidad. Topografía ondulada. Afluente de agua más cercano a 160 m aproximadamente. Poca influencia antrópica. |
|
JEAB |
Finca privada (PSA) |
Parte media de una loma, elevación de 250 m s. n. m., con cobertura de un bosque maduro, pero sin presencia de árboles de gran tamaño, con un dosel establecido, pero con espacios abiertos. Sotobosque denso con presencia de palmas. Topografía ondulada. Afluente de agua más cercano a 118 m aproximadamente. Camino principal hacia Drake a 163 m y limitando con un potrero a 78 m hacia el este. Poca influencia antrópica. |
|
JEAR |
Finca privada (PSA) |
Parte alta de una loma, elevación de 474 m s. n. m., con cobertura de un bosque maduro, pero algunos claros presentes por la caída de árboles, por lo que existen bastantes espacios. Sotobosque denso. Topografía fuertemente ondulada. No se determinaron afluentes de agua cercanos. Trocha de acceso a fincas (no transitado por vehículos) a 50 m. Poca influencia antrópica. |
|
CALE |
Finca privada (PSA) |
Parte media de una loma, elevación de 185 m s. n. m., dominada por una cobertura de bosque maduro, pero que no cierran en su totalidad el dosel. Sotobosque denso con muchas palmas. Topografía fuertemente ondulada. No se determinaron afluentes de agua cercanos. Poca influencia antrópica. |
|
HUJI |
Finca privada (PSA) |
Parte media de una loma, elevación de 286 m s. n. m., con cobertura de un bosque maduro, pero con presencia de espacios abiertos. Sotobosque denso con presencia de palmas. Topografía fuertemente ondulada. No se determinaron afluentes de agua importantes y cercanos. Camino principal hacia Drake a 530 m y del centro del poblado de Rancho Quemado a 850 m. Por las características topográficas. Influencia antrópica. |
|
TRUR |
Finca privada (PSA) |
Parte media de una loma, elevación 293 m s. n. m., con cobertura de bosque maduro, claros importantes en el dosel, sotobosque muy denso. La topografía es ondulada. Afluente de agua a 50 m aproximadamente. La grabadora se instaló a 50 m del borde del bosque. Poca influencia antrópica. |
|
OSMO |
Finca privada (PSA) |
Parte media de una loma, elevación 57 m s. n. m., con cobertura de bosque maduro y presencia de árboles grandes y dosel cerrado con algunos espacios de entrada de luz. Sotobosque abierto en algunos sectores. La topografía es ondulada. No se determinaron afluentes de agua importantes y cercanos. Poca influencia antrópica |
|
COR1 |
Parque nacional |
Parte alta de loma con topografía levemente ondulada, elevación 200 m s. n. m. Área de bosque secundario en estado avanzado de sucesión, con un dosel bien establecido y cerrado. Sotobosque denso. Se encuentra en el borde del Parque Nacional Corcovado, pero colinda con fincas de cobertura boscosa. Poca influencia antrópica. |
|
COR2 |
Parque nacional |
Sitio plano, elevación 217 m s. n. m. Área de bosque maduro, con un dosel bien establecido, pero con la presencia de claros. Sotobosque medianamente denso. Dentro del Parque Nacional Corcovado, a unos 430 m del límite. Afluente de agua como a 50 m. Poca influencia antrópica. |

Figura 1. Ubicación de los sitios de muestreo donde se colocaron las grabadoras en la Reserva Forestal Golfo Dulce, Osa, Costa Rica.
Figure 1. Location of the sampling sites where the recorders were placed in the Golfo Dulce Forest Reserve, Osa, Costa Rica.
Se colocó una grabadora Song Meter 4 (SM4; Wildlife Acoustics Inc.) en cada uno de los 11 sitios de muestreo a un mínimo de 100 m del borde del bosque (con excepción del sitio TRUR, que por sus caracteristicas la grabadora se ubicó a 50 m del borde de bosque) y a una altura de entre 1.8 a 2 m, con lo cual se evita árboles que podrían causar interferencia con los sonidos. Los audios fueron registrados en estéreo, en formato wav, 16 bits de resolución y con una tasa de muestreo de 32 kHz. Los dispositivos se programaron para grabar de manera continua en los bloques durante el amanecer (05:00-07:00) y el atardecer (17:00-18:00), así como grabaciones intermitentes de 5 minutos con intervalos de descanso de 10 minutos desde las 7:00 hasta las 16:50 horas. Este esquema de grabación se mantuvo durante 8 días consecutivos desde el 25 de mayo del 2021 hasta el 01 de junio del 2021.
2.2 Caracterización de sitios
En cada sitio se tomaron fotografías hemisféricas del dosel con un lente universal ojo de pez para celulares y fotografías de cuatro puntos alrededor de la parcela y se realizó una estimación del área basal utilizando un Cruiser´s Crutch (SEC, Inc.) en escala métrica.
El análisis de las fotografías se realizó con el progra.m.a Gap Light Analizer (GLA; versión 2.0). Se utilizaron los parámetros por defecto ajustando únicamente la opción Clear-Sky Transmission Coefficient y el umbral de acuerdo con la cantidad de brillo de cada fotografía hasta obtener un ajuste óptimo. Se calculó el porcentaje de apertura de sitio (apertura dosel), que indica cuánto espacio abierto existe en el sitio para la entrada directa de luz. Además, se calculó el índice de área foliar efectiva considerando un zenit de 0° a 60°.
Se delimitó un búfer de 500 m alrededor de la ubicación de cada grabadora y se calculó una serie de métricas de paisaje, utilizando como base el mapa de cobertura y uso del suelo del Atlas de Costa Rica 2014 (Cuadro 2). Estas variables fueron seleccionadas de acuerdo con su posible efecto sobre la biodiversidad, reportado en la literatura (Fahrig, 2003), y como una manera de caracterizar cada sitio de muestreo en función de su matriz circundante. El análisis se realizó en ArcGIS (ESRI, 2016) con la extensión Patch Analyst (Rempel et al., 2012).
Cuadro 2. Variables de paisaje utilizadas para caracterizar los sitios de muestreo, en un área circular de 500 m de radio alrededor de la grabadora.
Table 2. Landscape variables used to characterize the sampling sites, in a circular area of 500 m radius around the recorder.
|
Nombre |
Código |
Descripción |
|
Número de parches |
NumP |
Número total de parches de todas las clases. |
|
Porcentaje de bosque maduro |
Bosque |
Porcentaje de cobertura de bosque. |
|
Tamaño promedio de parches |
MPS |
Tamaño promedio de los parches (m2). |
|
Borde total |
TE |
Perímetro total de los parches (m). |
|
Promedio de índice de forma |
MSI |
Medida de la forma y grado de compactación medio del parche. Los valores van de 1 a infinito. Los valores bajos indican una forma compacta y los valores más altos, formas más irregulares. |
|
Apertura del dosel |
apertura_dosel |
Disminución de la densidad o espesura de la masa, para facilitar el crecimiento de los árboles que quedan en pie y la llegada de luz al suelo. |
|
Área basal |
area_basal |
Superficie total de la sección transversal de un tronco de un árbol, medida a una altura específica desde la base de este. |
|
Área foliar |
area_foliar |
Medida de la superficie total de las hojas de la vegetación. |
2.3 Cálculo de índices acústicos
Los índices TE, Ht, Hf, MAE y NDSI fueron calculados con base en las funciones descritas en el paquete seewave (Sueur et al., 2008). Los índices ADI, AEI, BIO y ACI se calcularon de acuerdo con las funciones del paquete soundecology (Villanueva-Rivera y Pijanowski, 2018), al modificar el ámbito de frecuencias empleado de 2 000 a 11 000 Hz, pues es más adecuado para cubrir la diversidad de sonidos en zonas tropicales (Pieretti, com. pers; Cuadro 3).
Cuadro 3. Descripción de los índices acústicos calculados en las fincas bajo pago por servicios ambientales en la Reserva Forestal Golfo Dulce (2022).
Table 3. Description of the acoustic indices calculated on farms under payment for environmental services in the Golfo Dulce Forest Reserve (2022).
|
Índice |
Descripción |
Categoría |
|
Total entropy (TE)/Entropía total |
Indicativo de la entropía en un segmento de audio a través del tiempo y frecuencias. Valores altos indican mayor actividad y diversidad acústica. Ámbito = [0, 1] (Towsey et al., 2014). |
Índices de entropía y diversidad acústica |
|
Temporal entropy (Ht)/Entropía temporal |
Indica la entropía entre cuadros de tiempo. Calcula la entropía de la suma de los valores de amplitud de cada cuadro de tiempo. Señales con muchas variaciones de amplitud tienden a valores cercanos a 1, mientras que señales acústicas planas o constantes, a valores cercanos a 0. Ámbito = [0, 1] (Towsey et al., 2014). |
Índices de entropía y diversidad acústica |
|
Spectral entropy (Hf)/Entropía espectral |
Indica la entropía entre bandas de frecuencia. Calcula la entropía de la suma de los valores de amplitud para cada banda de frecuencia. Señales con tonos puros, distribuidos en bandas de frecuencia muy compactas tienden a valores cercanos a 0, mientras que señales amplias, distribuidas en bandas de frecuencia amplias tienden a valores cercanos a 1. Ámbito = [0, 1] (Towsey et al., 2014). |
Índices de entropía y diversidad acústica |
|
Acoustic complexity Index (ACI)/Índice de complejidad acústica |
Medida que cuantifica la variación en la amplitud a través del tiempo en el espectro. Valores altos se relacionan con mayor actividad acústica. Modulaciones rápidas aumentan el valor del ACI. Ámbito = [0, +] (Pieretti et al., 2011). |
Índices de entropía y diversidad acústica |
|
Acoustic diversity index (ADI)/Índice de diversidad acústica |
Estima la diversidad acústica en el espectro de frecuencias. Es el resultado del índice de diversidad de Shannon aplicado en la proporción de señales que sobrepasan un umbral de amplitud en cada banda de frecuencia. Valores altos indican que las señales acústicas están distribuidas por todo el espectro. Valores bajos, que las señales acústicas están concentradas en pocas bandas de frecuencia. Ámbito = [0, 1] (Villanueva-Rivera et al., 2011). |
Índices de entropía y diversidad acústica |
|
Acoustic evenness index (AEI)/Índice de equidad acústica |
Es una medida de equidad en la distribución de energía a través del espectro. Es el resultado del índice de equidad de Gini aplicado sobre la proporción de señales que sobrepasan un umbral de amplitud en cada banda de frecuencias. Valores cercanos a 0 representan alta equidad; es decir, que las señales acústicas están distribuidas equitativamente en todo el espectro de frecuencias. Ámbito = [0, 1] (Villanueva-Rivera et al., 2011). |
Índices de entropía y diversidad acústica |
|
Bioacoustic Index (BIO)/Índice bioacústico |
Indica la energía acústica ocupada en las bandas de frecuencia entre 2 000 y 11 000 Hz (asociadas a biofonías). Valores altos reflejan alta actividad acústica en dichas bandas de frecuencia. Ámbito = [0, +] (Boelman et al., 2007). |
Índices de resumen de energía acústica |
|
Median Amplitude Envelope (MAE)/Mediana de la amplitud envolvente |
Indica la mediana de la envolvente de amplitud normalizada por el valor máximo. Valores cercanos a 1 indican ambientes más ruidosos. Ámbito = [0, 1] (Depraetere et al., 2012). |
Índices de resumen de energía acústica |
|
Normalized difference soundscape index (NDSI)/Índice de paisaje sonoro de diferencia normalizada |
Es una medida normalizada de relación entre la energía contenida en las bandas de frecuencia usualmente ocupadas por sonidos biológicos (2 000-8 000 Hz) y las ocupadas por sonidos antrópicos (1 000-2 000 Hz). +1 representa la energía concentrada completamente en las frecuencias 2 000-8 000 Hz; -1 representa energía concentrada solo en las bandas antrópicas de 1 000-2 000. Ámbito = [-1, +1] (Kasten et al., 2012). |
Índices de resumen de energía acústica |
|
Mean Sound Pressure Level (SPLm)/Nivel de presión de sonido promedio |
Es un indicativo del nivel de presión de sonido promedio en el lugar, en dB. Valores altos reflejan ambientes más ruidosos. Ámbito = [0, +]. |
Índices de resumen de energía acústica |
Se revisaron manualmente 11 601.5 minutos de grabación provenientes de 6 grabadoras. Se etiquetó cada minuto correspondiente a lluvia fuerte, presencia de chicharras o un archivo limpio (sin los anteriores). Para cada minuto se calcularon datos estadísticos del espectro (centroide, intercuantil, tercer cuantil, asimetría (skewness) y curtosis), la dominancia de cada banda de frecuencia de 1 kHz desde los 0 hasta los 11 kHz, la frecuencia dominante promedio y la mediana del nivel de ruido promedio siguiendo el procedimiento de Towsey (2013).
Estos datos fueron utilizados para construir el modelo Random Forest (Breiman, 2001) con el objetivo de clasificar los minutos de audio en las categorías de Lluvia, Chicharra y Limpio. El modelo se aplicó sobre todos los audios. Los minutos de audio clasificados como Lluvia y Chicharra fueron excluidos de posteriores análisis. Para reducir la ocurrencia de valores atípicos producidos por eventos sonoros azarosos, se eliminaron los valores extremos definidos como los valores 1,5 veces mayores y menores al ámbito del intercuantil (3.er cuantil-1.er cuantil).
2.5.1 Comparación de índices por conglomerados
Los índices fueron comparados en conglomerados para analizar las diferencias en el paisaje sonoro, no a nivel individual, sino como conjunto de sitios que comparte características de paisaje. Para este análisis se implementó un modelo de clasificación mediante el análisis de conglomerados por el método K-means (Hartigan y Wong, 1979). Las variables seleccionadas para la clasificación se describen en el Cuadro 1. Se aplicó un modelo mixto generalizado para analizar la respuesta de cada índice acústico de acuerdo con el conglomerado. Se utilizó la mediana por hora y por sitio de los valores de cada índice como variable respuesta; el conglomerado, la hora del día y la interacción entre ambos como efectos fijos, así como el sitio como efecto aleatorio.
Adicionalmente, cada modelo incluyó una estructura de covarianza AR1 para considerar la autocorrelación entre horas consecutivas del día. Los diferentes ajustes de distribución de los residuales fueron evaluados mediante simulaciones por medio del paquete DHARMa (Hartig, 2022). Como resultado, los modelos para los índices Ht, Hf, TE, AEI, MAE y NDSI fueron construidos según una estructura de distribución de residuales betas. Por su parte, los índices BIO y ACI se modelaron con una distribución Gama, el índice ADI con una distribución gaussiana.
2.5.2 Influencia de los índices en la clasificación
Se implementó un modelo de Random Forest (Breiman, 2001) para clasificar los conglomerados de acuerdo con los índices acústicos e identificar los índices más influyentes en esta clasificación. Esto permitirá analizar cuáles presentan mayor diferencia entre un conglomerado y otro. El modelo se construyó utilizando la mediana por hora y día de cada índice para todos los sitios. Se descartaron los valores extremos.
Para estimar la importancia de cada variable en el modelo de clasificación, se utilizó la función varImp del paquete caret (Kuhn, 2008), la cual, para cada árbol del modelo, registra el valor de precisión de la predicción en la parte de los datos antes de procesarlos (out-of-bag), se repite luego de permutar cada variable predictora y la diferencia entre las dos precisiones es promediada en todos los árboles y normalizada por el error estándar.
2.5.3 Comparación de índices por sitio
Los sitios fueron analizados y comparados de acuerdo con cada uno de los índices acústicos seleccionados. Se aplicó un modelo lineal generalizado (Brooks et al., 2017) utilizando la mediana por hora y sitio de los valores de cada índice acústico como variable respuesta. El sitio y la hora se consideraron variables explicativas. Los valores de NDSI fueron transformados a valores entre 0-1 según la fórmula: (NDSI+1)/2. Se evaluaron diferentes ajustes de distribución de los residuales mediante simulaciones utilizando el paquete DHARMa (Hartig, 2022) para seleccionar el modelo que mejor se concordara con la distribución de los datos. Como resultado, los modelos para los índices AEI, TE, Ht, Hf, MAE y NDSI se ajustaron utilizando una distribución beta; los índices ADI, BIO, SPLm con una distribución gaussiana y ACI con una distribución gaussiana con un enlace logarítmico.
2.5.4 Detección acústica de especies
Se seleccionaron las especies trogón vientribermejo (Trogon bairdii) y el tororoi pechilistado (Hylopezus perspicillatus) como modelos para la detección automatizada en los audios de los sitios de estudio. Estas especies fueron seleccionadas por sus requerimientos de hábitat (como dependencia de bosques maduros y características del sotobosque), así como por sus vocalizaciones fácilmente reconocibles.
En el caso de T. bairdii, al ser una especie endémica de Costa Rica y Panamá, es relativamente común en las tierras bajas del Pacífico Sur de Costa Rica donde habita en bosques húmedos (Garrigues y Dean, 2014). Por su parte, H. perspicillatus es una especie común del Pacífico Sur de Costa Rica, habita en bosques húmedos maduros donde prefiere ambientes con un sotobosque abierto (Garrigues y Dean, 2014).
Para identificar los parámetros de delimitación de las vocalizaciones de ambas especies se analizaron los audios disponibles en la librería digital de Xeno-Canto (www.xeno-canto.org) grabados dentro del territorio costarricense. Se utilizó la herramienta de selección en el software Raven Pro 1.6.3 (K. Lisa Yang Center for Conservation Bioacoustics, 2022) para delimitar y obtener los valores de Frecuencia máxima, Frecuencia mínima, Duración y Máximo intervalo entre sílabas de cada vocalización; estos fueron empleados como parámetros delimitantes.
Las vocalizaciones se detectaron con el Kaleidoscope Pro (Wildlife acoustics Inc.). Se creó un clasificador avanzado para cada especie, el cual consiste en ajustar los parámetros de detección con base en un conjunto de audios con señales de las especies de interés (Wildlife acoustics Inc., 2023). El clasificador se aplicó en cada uno de los sitios de estudio utilizando los audios de las fechas comprendidas entre el 25 de mayo y el 1 de junio del 2020, para una muestra total de 8 días. Las detecciones identificadas fueron revisadas manualmente para corroborar todas las vocalizaciones identificadas. El porcentaje de detecciones fue calculado con base en la cantidad de días donde se detectaron vocalizaciones de la especie de interés, dividido entre la cantidad total de días de muestreo (8 días).
Los sitios JEAB, HUJI, TRUR y OSMO presentaron fragmentación, siendo el primero el más fragmentado, con 8 parches. Por su parte, los demás sitios (CALE, COR1, COR2, CRRI, HEMO, JEAR y JUCU) estuvieron integrados por un solo parche boscoso. Esto se reflejó también en las métricas de tamaño promedio de parche (MPS) y el borde total (TE; Figura 2). Las variables de estructura vegetal, a su vez, mostraron una tendencia menos marcada. COR1 y JEAR presentaron la mayor apertura de dosel, mientras que JUCU, la menor. HUJI presentó una mayor área foliar y COR1, la menor. Finalmente, COR2, HEMO y JUCU obtuvieron la mayor área basal y TRUR, CALE, JEAR y OSMO, la menor (Figura 2).
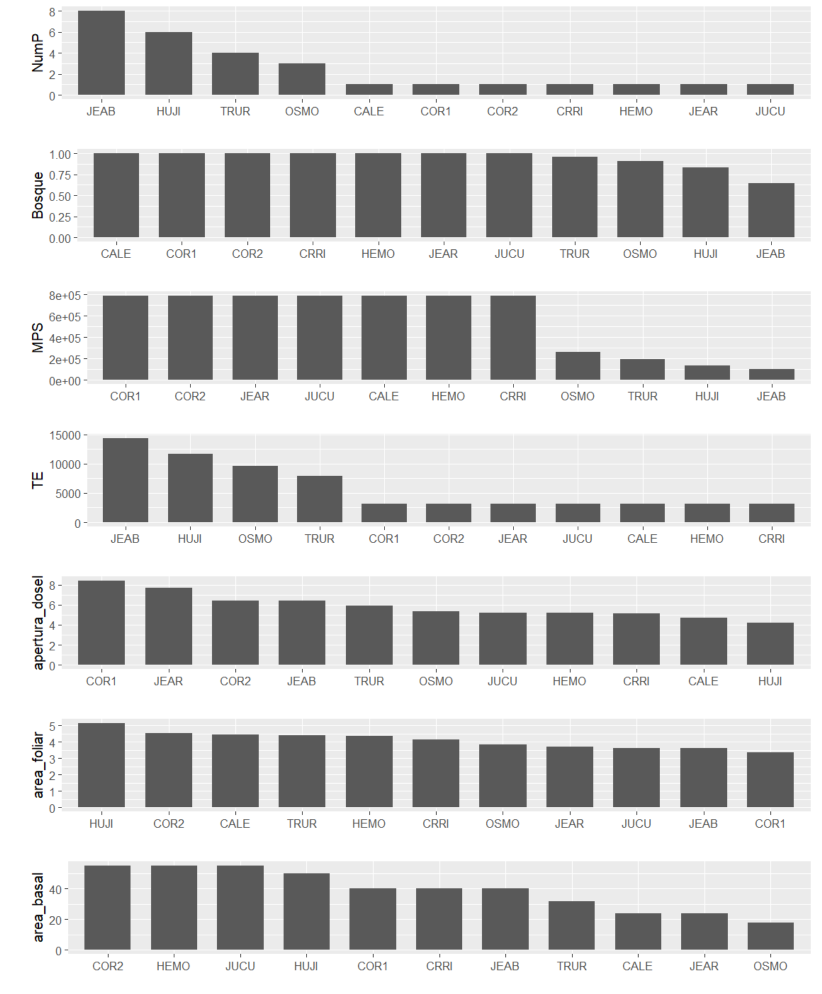
Figura 2. Caracterización de sitios de muestreo de acuerdo con las variables de paisaje y estructura vegetal en la Reserva Forestal Golfo Dulce.
Figure 2. Characterization of sampling sites according to landscape variables and vegetation structure in the Golfo Dulce forest reserve.
El análisis de conglomerados agrupa los sitios JEAB, OSMO, TRUR y HUJI en el cluster2, y a los demás en el cluster1 (Figura 3).
El desempeño de la agrupación fue de 0.43 según el índice de ancho promedio de silueta (Average silhouette width). El ámbito de este índice va de -1 a 1, donde 1 es una agrupación perfecta de los sitios, valores cercanos a 0 indican un mal desempeño de la agrupación y lo negativos, que los sitios fueron agrupados en un conglomerado incorrecto.
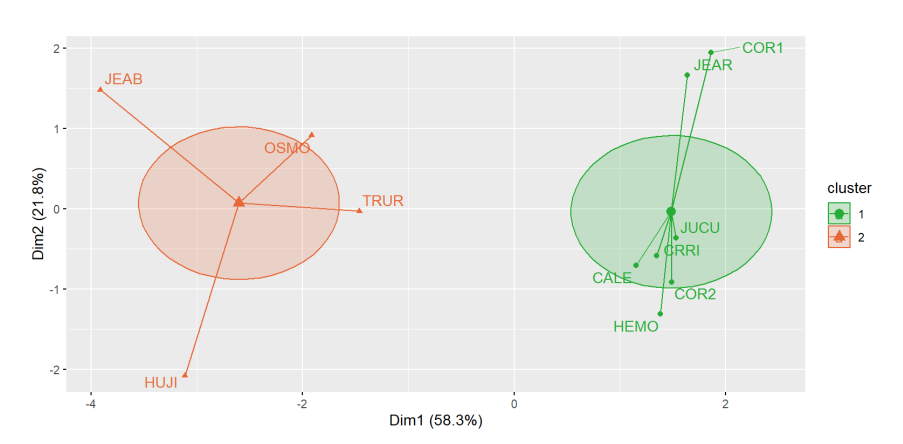
Figura 3. Separación de sitios mediante el método K-means de acuerdo con las variables de paisaje.
Figure 3. Separation of sites using the K-means method according to landscape variables.
El cluster1 se caracterizó por contener sitios con una mayor proporción de bosque, mayor tamaño promedio de parches, menor número de parches, menor borde y menor índice de forma de parche. A su vez, el cluster2 se identificó por una menor proporción de bosque, menor tamaño de parches promedio y mayor índice de forma, número de parches y borde total (Cuadro 4). Las variables más importantes en la clasificación de los clústeres son el porcentaje de bosque y el número de parches (Cuadro 4).
Cuadro 4. Promedio del valor de variables de vegetación y de paisaje con su error estándar para cada conglomerado de sitios.
Table 4. Average value of vegetation and landscape variables with their standard error for each cluster of sites.
|
Variable |
Cluster 1 |
Cluster 2 |
Valor de importancia (%) |
|
Bosque |
1.00 ± 0 |
0.837 ± 0.21 |
100.00 |
|
NumP |
1.00 ± 0 |
5.25 ± 3.53 |
85.40 |
|
MSI |
1.00 ± 0 |
2.08 ± 0.33 |
84.72 |
|
TE |
3 143.44 ± 0.17 |
10 887.28 ± 4 381.99 |
75.88 |
|
MPS |
786 216.86 ± 85.06 |
171 982.97 ± 115 567.80 |
71.32 |
|
Apertura_dosel |
6.12 ± 1.32 |
5.46 ± 1.52 |
4.96 |
|
Área_foliar |
4.01 ± 0.42 |
4.23 ± 1.07 |
3.00 |
|
Área_basal |
41.86 ± 12.87 |
35 ± 21.51 |
0.00 |
3.3 Variación de los índices acústicos por conglomerados
El índice ACI fue el único índice acústico evaluado que presentó una variación significativa entre los conglomerados. Todos los demás índices acústicos presentaron variación durante el día, pero siguiendo la misma tendencia en ambos conglomerados sin una diferencia estadísticamente significativa (Figura 4).
En el caso del índice de complejidad acústica (ACI), el cluster2 presentó valores significativamente mayores al cluster1 (Figura 4), en especial durante las horas de la mañana (5:00-8:00). Por su parte, de manera general el cluster2 mostró una tendencia a valores mayores de diversidad y entropía acústica (ADI, AEI, Hf, TE), con excepción de la entropía temporal que fue mayor en el cluster1.
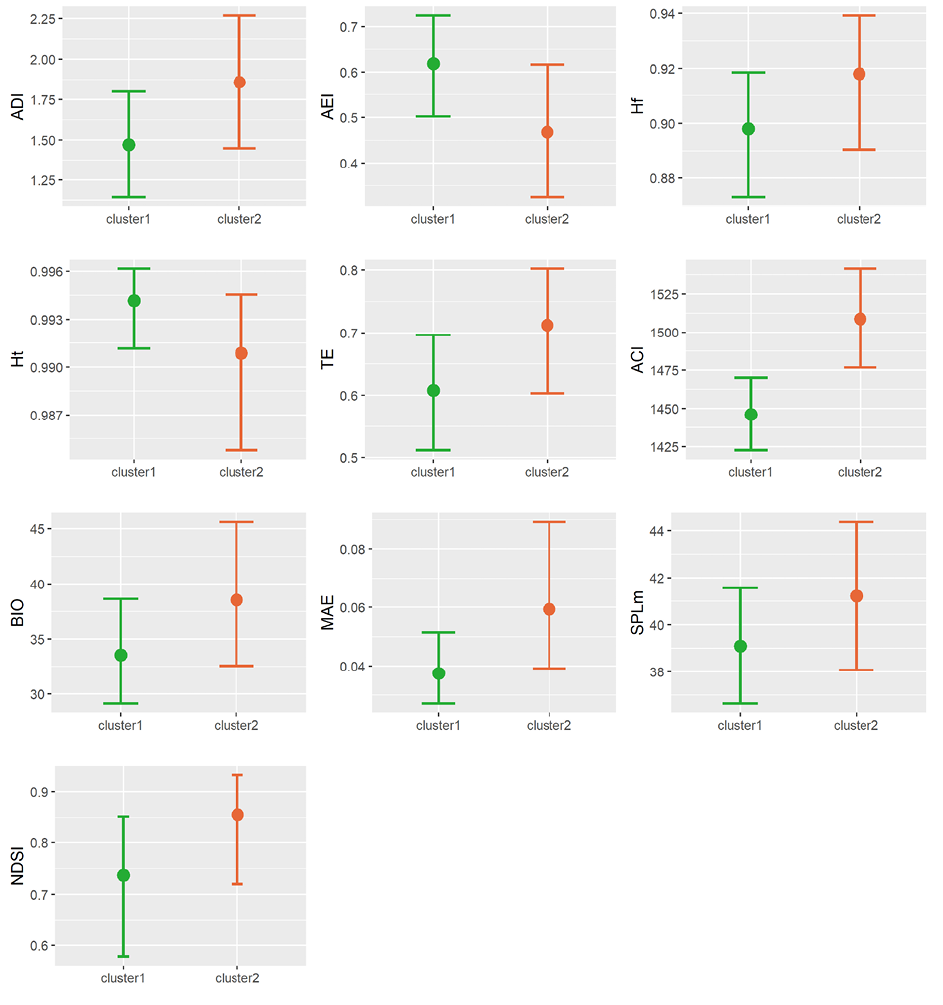
Figura 4. Comparación entre conglomerados de valores de índices acústicos.
Figure 4. Comparison between clusters of acoustic indices values.
A pesar de no obtener una diferencia estadísticamente significativa, la tendencia observada es que el cluster2 (más fragmentado) presenta valores mayores en los índices de resumen de energía (índices BIO, MAE, SPLm y NDSI; Figura 4), esto indica una mayor energía acústica contenida en este conjunto de sitios. En el caso del índice NDSI, la proporción de energía en las bandas de frecuencia relacionadas con biofonías con respecto a antrofonías es mayor en el cluster2.
3.3.1 Importancia de índices acústicos en la clasificación de los conglomerados
El modelo de clasificación Random Forest clasificó ambos conglomerados de acuerdo con los valores de los índices acústicos con un 93.7 % de precisión. Los índices con mayor valor de importancia en la clasificación fueron SPLm y MAE (Figura 5).
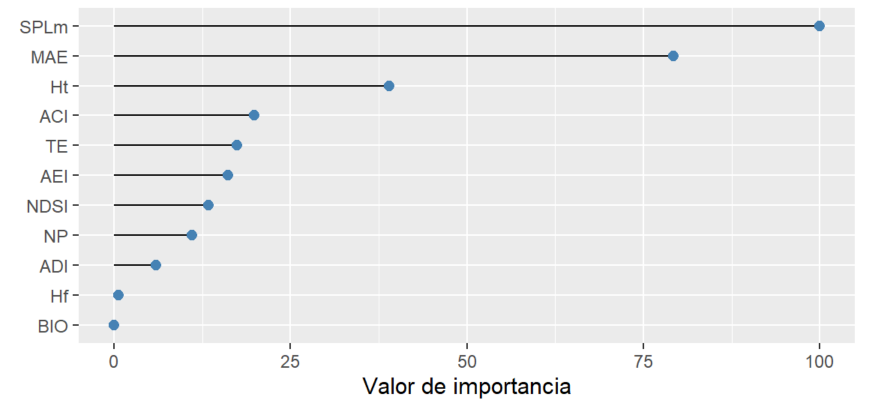
Figura 5. Valor de importancia de cada índice acústico según el modelo de clasificación Random Forest por conglomerado.
Figure 5. Importance value of each acoustic index in the Random Forest classification model by cluster.
Los índices que mayor influyen en la diferenciación entre ambos conglomerados son aquellos que cuantifican la energía acústica en el ambiente (SPLm y MAE); es decir, el mayor determinante entre los sitios agrupados en el cluster1 y el cluster 2 es la cantidad de energía acústica que contiene el espectro acústico.
3.4 Comparación de índices por sitios
Los índices acústicos variaron de acuerdo con el sitio evaluado. No obstante, cada uno presentó una tendencia diferente en su variación con respecto a los sitios (Figura 6).
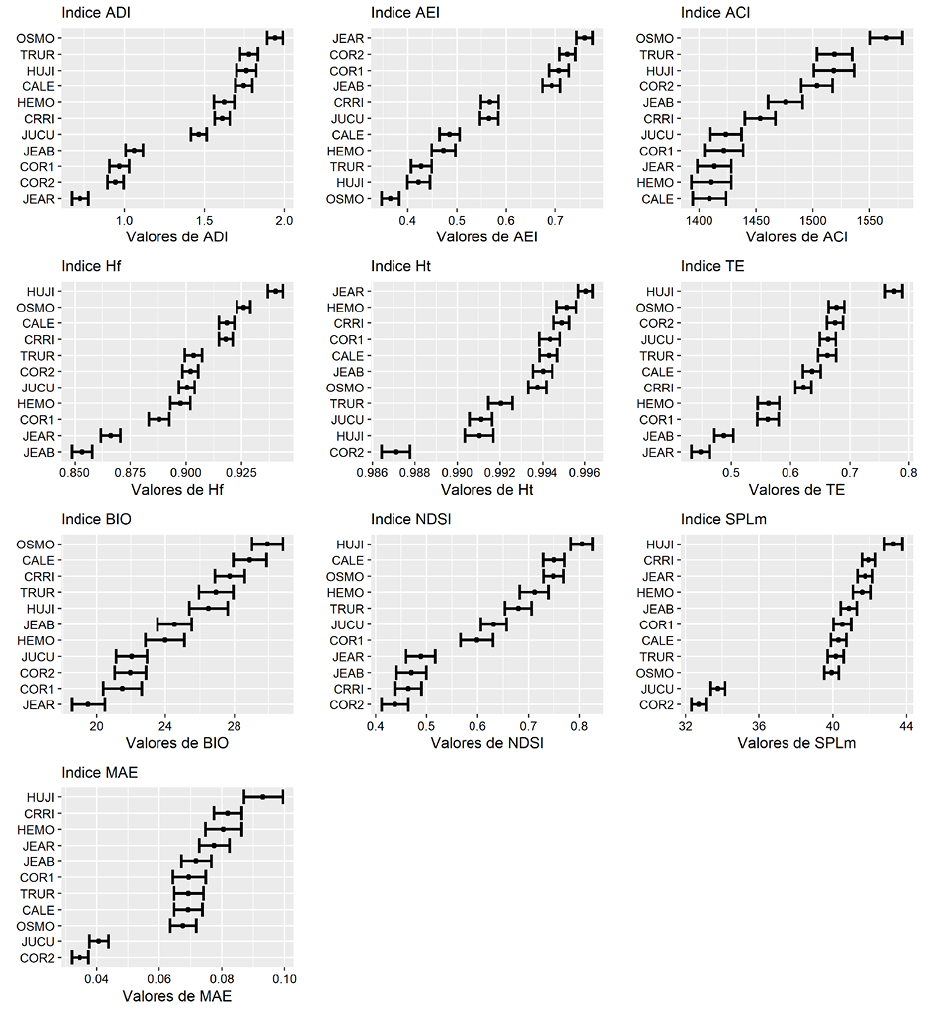
Figura 6. Comparación de los valores promedio de los índices acústicos por sitio.
Figure 6. Comparison of acoustic indices values by site.
El sitio con mayor ADI y menor AEI (por ende, mayor equidad acústica) fue OSMO, seguido por TRUR. Por otra parte, los sitios que presentan los valores más bajos de diversidad y equidad acústica son JEAR seguido por COR1 y COR2 (Figura 6).
OSMO destaca como el sitio con mayor complejidad acústica (índice ACI). COR2, a pesar de ser uno de los sitios más silenciosos y con poca energía acústica (bajo SPLm y MAE; Figura 6), fue uno de lo que obtuvo mayor ACI.
La mayor Hf fue registrada en el sitio HUJI, seguido de OSMO, CRRI y CALE (Figura 6). Los valores más bajos de Hf pertenecen a JEAR y JEAB, lo cual indica una menor variación en las frecuencias de sonidos que se encuentran en estos sitios.
El sitio con mayor entropía temporal (Ht) fue JEAR, seguido de cerca por CALE, COR1, CRRI, HEMO, JEAB y OSMO con valores muy similares. COR2 se convirtió en el sitio con menor entropía temporal, a pesar de estar muy cerca, espacialmente, al sitio COR1. Seguido de este se encuentra HUJI como el segundo más bajo (Figura 6).
La entropía total (TE) se comportó de manera similar a la de frecuencias (Hf; Figura 6). El sitio con valores más altos fue HUJI, mientras que JEAB y JEAR presentan los menores valores de entropía, lo cual los relaciona con los sitios con menor variación en la energía acústica. Ambos sitios (JEAB y JEAR) son cercanos y mantuvieron la misma tendencia con respecto a este índice.
Ahora bien, el sitio HUJI destaca por ser el de mayor energía acústica, la cual está concentrada en las bandas de frecuencia comúnmente utilizadas por biofonías (2 000-8 000 Hz), como lo demuestran los índices BIO y NDSI. Además, CALE, OSMO, CRRI y TRUR obtuvieron valores muy similares y relativamente altos del índice BIO. La mayor energía se concentra en horas de la mañana (Figura 12).
Los sitios menos ruidosos (más silenciosos) son COR2 y JUCU, reflejado en ambos índices SPLm y MAE. Este resultado se evidencia también en los valores del índice BIO, estos sitios muestran una baja actividad acústica de sonidos biológicos.
3.5 Detección acústica de especies
Se registró un total de 1 909 vocalizaciones para Trogon bairdii y 423 para Hylopezus perspicillatus. La especie T. bairdii se detectó en todos los sitios con excepción de JEAR, la probabilidad de detección diaria de esta especie fue de 0.5 o mayor en la mayoría de los sitios (6 de 11) y en el caso del sitio OSMO fue detectado durante todos los días evaluados (Figura 7).
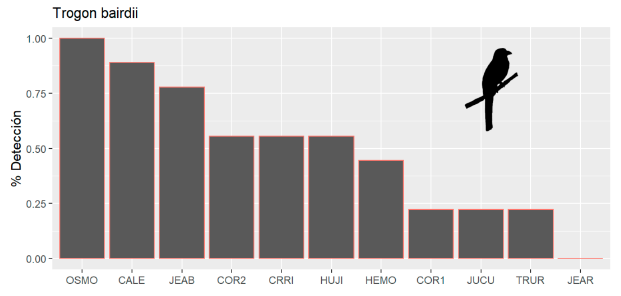
Figura 7. Porcentaje de días en los que se detectó la presencia de Trogon bairdii en cada sitio de estudio.
Figure 7. Percentage of days in which the presence of Trogon bairdii was detected in each study site.
La detección de Hylopezus perspicillatus fue más baja que la de T. bairdii. Esta especie se encontró únicamente en 3 de los 11 sitios. El lugar con mayor cantidad de detecciones fue COR1, con un total de 417 detecciones y una probabilidad de detección diaria de 0.55. Los otros sitios donde se encontró fueron COR2 (7 registros) y HEMO (2 registros), para una probabilidad de detección diaria de 0.22 en ambos (Figura 8).

Figura 8. Porcentaje de días en los que se registraron vocalizaciones de Hylopezus perspicillatus en cada sitio de estudio.
Figure 8. Percentage of days in which Hylopezus perspicillatus vocalizations were detected at each study site.
Las variables asociadas a la fragmentación del bosque resultaron ser las más relevantes en la caracterización de los sitios. Otras, como la apertura del dosel, el área foliar y el área basal, no resultaron determinantes, probablemente por contener una cobertura vegetal similar. En el caso del área basal, quizás influyó el método de cuantificación, en el que se determinó a través de parcelas de radio variable, en donde, a pesar de valorar la ocupación, esta no refleja la distribución de dicha ocupación dentro de la estructura horizontal del bosque (Keene y Barlow, 2019). Por lo tanto, sitios con árboles de gran porte, pero menos densos, pueden equipararse en valor de ocupación con áreas de bosque más densas, pero con árboles de menor tamaño diamétrico.
Los resultados del análisis acústico de los conglomerados presentaron mucha variabilidad, tanto entre sitios como entre los conglomerados en análisis. No obstante, se encontró que los sitios más fragmentados tienen una tendencia a presentar mayor actividad, energía y diversidad acústica que aquellos menos fragmentados. Este patrón posiblemente se deba a los efectos de borde y ecotonos en los sitios fragmentados, así como a un alto componente de bosque en la matriz circundante.
El análisis de los sitios en conglomerados fue poco concluyente porque las diferencias en la mayoría de los valores de los índices acústicos entre conglomerados no son estadísticamente significativas. Esto puede deberse a la similitud entre los sitios de estudio. A pesar de las diferencias en la cantidad de parches, el porcentaje de bosque y las variables de estructura vegetal como área foliar, apertura de dosel y área basal, todos los sitios cuentan con un alto porcentaje de bosque (mayor a 65 %). Además, en muchos casos los sitios mantienen conexión con áreas de bosque más extensas e incluso los bosques de diferentes sitios se conectan entre sí.
Además, la cercanía y similitud en las características de los sitios dificulta observar patrones contrastantes (Vega-Hidalgo et al., 2021). En áreas poco contrastantes se recomienda mejorar la caracterización de los sitios, considerando otros métodos más detallados para determinar la estructura horizontal e incluir aspectos de estructura vertical y de composición de especies vegetales, de forma que facilite el evaluar aspectos específicos de los sitios, más allá de variables generales.
A pesar de las similitudes, el índice ACI fue lo suficientemente sensible para mostrar diferencias significativas, desde un punto de vista estadístico, entre los conglomerados. La tendencia observada sugiere que los sitios con una matriz fragmentada (con la excepción de COR2, el cual es un sitio no fragmentado) presentan valores más altos de ACI. Esto concuerda con otros estudios en zonas tropicales donde los bosques secundarios presentan una mayor actividad acústica dada la influencia de efectos de ecotono, o bien la presencia de especies dominantes (Fahrig et al., 2019).
Adicionalmente, se ha encontrado que los sitios con presencia humana suelen tener valores de ACI más altos de lo esperado (Fairbrass et al., 2017; Mitchell et al., 2020). Ante esto, la diferencia entre conglomerados quizá se deba a la alteración humana como influencia de carreteras o cercanía a poblados. En los valores altos de ACI en COR2 puede deberse a un factor diferente: los audios de este sitio registraron sonidos atípicos, pulsaciones rápidas de gran energía (tipo “click”) que aparecen esporádicamente en la grabación (ver Anexo 1). Se infiere que estos sonidos pueden deberse a un mal funcionamiento de la grabadora o a un deterioro de la cubierta protectora de los micrófonos, propiciando la intromisión de ruidos por contacto físico del micrófono con algún componente ambiental. Estos sonidos incrementarían los valores de AIC debido a su naturaleza de pulsaciones rápidas con gran intensidad y explicaría el patrón observado en este lugar.
Los índices SPLm y MAE fueron los más importante en la clasificación de conglomerados de acuerdo con los índices acústicos. Los valores más bajos de SPLm y MAE se asocian principalmente con sitios conservados, siendo estos los más silenciosos (Gonçalves et al., 2021). Estas diferencias en la energía acústica pueden deberse a la estructura y composición de la vegetación y su relación con la propagación del sonido (Slabbekoorn, 2004), o bien a la influencia de sonidos antrópicos (Fairbrass et al., 2017; Mitchell et al., 2020). Por ejemplo, las barreras físicas pueden interferir en la propagación del sonido y atenuar sonidos externos (Yip et al., 2017), por lo que los sitios con una estructura de vegetación densa o una topografía capaz de bloquear los sonidos (como valles, por ejemplo) pueden propiciar ambientes más silenciosos. De igual forma, los sonidos antrópicos en los sitios alterados pueden incrementar los valores de energía acústica registrado por los índices.
Los sitios evaluados presentan considerable fluctuación en los valores de los índices acústicos. No obstante, al comparar las características de paisaje correspondiente a cada sitio con los valores de índices acústicos, se encuentra que los sitios fragmentados parecen registrar mayor diversidad acústica y entropía de frecuencias (índices: ADI, AEI, Hf y TE) que aquellos con una cobertura total de bosque. Esto también sucede con el índice BIO, donde se evidencia mayor energía acústica en los sitios con mayor cantidad de parches o presentan una estructura vegetal característica de bosques más abiertos (p. ej., HUJI, CALE, OSMO); es decir, son más activos biológicamente. Estos sitios también obtuvieron valores altos de diversidad acústica (ADI), por lo que se podría inferir que mantienen una mayor diversidad o abundancia de especies activas desde el punto de vista acústico (Villanueva-Rivera et al., 2011). Por su parte, los sitios con menor energía acústica (JUCU y COR2) se componen solo de bosque maduro y presentan la mayor área basal, características de bosques conservados.
Una explicación es que los bosques más abiertos o secundarios suelen presentar mayor actividad biológica y una composición diferente de especies (Valente y Betts, 2019). Ahora bien, aquellos con una matriz fragmentada muestran ecotonos donde convergen especies de distintos ambientes, por lo que resulta común encontrar mayor diversidad en sitios alterados que en bosques primarios (Carrara et al., 2015; Kark, 2013). Adicionalmente, la composición de especies se ve influida por el efecto de borde (Fahrig et al., 2019). Las especies generalistas resultan favorecidas por el ecotono, mientras que un efecto contrario sucede con las especialistas (Rosli et al., 2018). Por lo tanto, las especies vocales de los hábitats alterados podrían ser, en su mayoría, comunes y abundantes que incrementan la actividad acústica del sitio.
Además de esto, los ambientes fragmentados pueden mantener altos niveles de diversidad cuando se encuentran inmersos en una matriz favorable; por ejemplo, de bosque (Fahrig et al., 2019; Bhakti et al., 2018). Este es el caso de los sitios de estudio, donde si bien se presentan diferencias en la cantidad de parches, todos cuentan con un alto porcentaje de bosque y, en muchos casos, mantienen conexión con áreas de bosque más extensas. La cercanía y similitud en las características de los sitios dificulta observar patrones contrastantes (Vega-Hidalgo et al., 2021).
No se puede ignorar que las diferencias entre sitios se podrían deber a características particulares o locales de cada uno y no a una tendencia general de acuerdo con el tipo de bosque o matriz. Los datos e inferencias fueron obtenidos de una sola grabadora por cada sitio, siendo susceptible a sesgos en los valores de los índices debido a eventos azarosos localizados. Para evitar este sesgo, es recomendable asegurar una muestra representativa, tanto temporal como espacialmente, de todos los sitios (Bradfer‐Lawrence et al., 2019).
La especie H. perspicillatus resultó ser más sensible que T. bairdii a las diferencias en el paisaje de los sitios estudiados. La segunda, a pesar de ser una especie que depende de bosques maduros, puede y suele aprovechar los bordes de bosque para alimentarse de frutos (Schulze y Riedl, 2008), por lo que comúnmente hace uso de áreas alteradas durante ciertos periodos. Por el contrario, la primera es una especie sensible a la degradación del hábitat, presenta hábitos elusivos y huidizos (Carantón-Ayala y Certuche-Cubillos, 2010; Garrigues y Dean, 2014; Renjifo y Amaya-Villarreal, 2017) y suele encontrarse en bosques maduros con un sotobosque abierto (Garrigues y Dean, 2014). Los resultados obtenidos muestran correctamente estos patrones.
A su vez, se resalta la utilidad del monitoreo acústico-pasivo para el estudio de estas dos especies, así como también su importancia en la evaluación de ecosistemas de acuerdo con la presencia de especies clave. Esta metodología ha sido implementada en diversos estudios para mantener un monitoreo biológico sobre especies de interés con muy buenos resultados (Das et al., 2020).
En conclusión, las características del paisaje evaluado mediante índices acústicos presentan patrones que se asocian con características del grado de conservación de un sitio; por ejemplo, la cantidad de energía acústica es mayor en sitios con características de bosques alterados, mientras que aquellos más conservados presentan patrones sonoros con menor energía acústica. Sin embargo, el uso solo de índices acústicos no resultó un elemento realmente diferenciador de la calidad del ecosistema en las fincas evaluadas, sobre todo en sitios no tan contrastantes en cuanto a la cobertura vegetal presente. Estos resultados son una de las primeras aproximaciones al uso del monitoreo acústico como herramienta para incorporar el componente de biodiversidad en el esquema de pago por servicios ambientales.
Históricamente, la incorporación de biodiversidad en PSA ha sido un reto debido a la dificultad para definir este concepto (Ezzine de Blas et al., 2011). Si bien el monitoreo de especies clave resulta menos complejo para establecer indicadores y variables de evaluación de corto a mediano y largo plazo, se puede estar descuidando otros componentes de la biodiversidad (Ezzine de Blas et al., 2011).
Se insta a otros grupos investigadores a que profundicen en la evaluación de estos y otros índices acústicos como indicadores del estado de conservación de ecosistemas o para monitoreos de biodiversidad. Estudios recientes enfocados en relacionar los índices acústicos con componentes de la biodiversidad han encontrados resultados promisorios (Alcocer et al. 2022; Bradfer-Lawrence et al. 2020, Dröge et al., 2021). Por lo tanto, es posible que las limitaciones metodológicas implementadas en este trabajo dificultaran encontrar los patrones esperados.
Otra recomendación es mejorar el diseño de muestreo, tratando de disminuir la variabilidad de los sitios, aumentando la cantidad de grabadoras por finca y mejorando la representatividad de cada sitio, tanto espacial como temporalmente. Estas adaptaciones metodológicas pueden conllevar a mejoras e inferencias más robustas en la interpretación de los resultados (Metcalf et al., 2021). Además, se deben evaluar otros índices acústicos no considerados en este estudio.
Actualmente, la cantidad de índices acústicos asciende cada vez más, algunos son aplicados en ambientes marinos (Minello et al., 2021), otros se basan en algoritmos de inteligencia artificial (Nieto-Mora et al. 2023) y otros han sido más explorados (Alcocer et al., 2022). Sin embargo, es un hecho que se necesita una mayor evidencia para comprender su comportamiento y aplicabilidad en la evaluación de los componentes de la biodiversidad.
Finalmente, el monitoreo de especies mediante la detección de sus vocalizaciones resultó ser una herramienta promisoria, la cual podría complementar evaluaciones de la calidad de los bosques y podría ser relevante en las evaluaciones y seguimientos en procesos de PSA. Sin embargo, se requiere de mayores estudios que asocien ciertas condiciones de los ecosistemas con especies clave presentes y que sean detectables con facilidad durante muestreos acústicos. El asocio de las características del ecosistema (favorables o desfavorables) con especies determinadas, podría facilitar procesos de evaluación y reducir los efectos que tienen los índices en zonas con alta presencia de sonidos no biológicos, en especial antrópicos.
Se extiende un agradecimiento a las personas de las fincas que permitieron el ingreso a sus propiedades; así como al personal del Área de Conservación de Osa por su ayuda en campo y en el trámite del permiso de investigación (resolución N.º SINAC-ACOSA-DASP-PI-R-023-2021). También a Marilyn Ureña y Gerardo Velado, quienes colaboraron en la revisión y etiquetado de audios para identificar presencia de lluvia.
El manuscrito deriva de la realización del proyecto de la Universidad Nacional titulado: “Creación de capacidades institucionales para el monitoreo acústico en fincas que forman parte del proceso de desarrollo de un sistema de pago de servicios ambientales en biodiversidad”, número SIA: 0258-19. Este se realizó para cumplir con los objetivos del convenio 07-2018 entre la Universidad Nacional y el FONAFIFO. Finalmente, se extiende un agradecimiento a las personas revisoras anónimas por sus comentarios y aportes para mejorar este trabajo.
6. Ética y conflicto de interés
Las personas autoras declaran que han cumplido a cabalidad con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en la producción del manuscrito; que no hay conflictos de intereses de ningún tipo; que todas las fuentes financieras se mencionan de forma completa y clara en la sección de agradecimientos y que están totalmente de acuerdo con la versión final editada del artículo.
Alcocer, I., Lima, H., Sugai, L. S. M. y Llusia, D. (2022). Acoustic indices as proxies for biodiversity: a meta‐analysis. Biological Reviews, 97(6), 2209-2236. https://doi.org/10.1111/brv.12890
Bermúdez-Rojas, T. y Obando-Acuña, V. (2021). Biodiversidad en cifras: avances en el conocimiento de especies en Costa Rica. Biocenosis, 32(2), 51-58. https://doi.org/10.22458/rb.v32i2.3899
Bhakti, T., Goulart, F., de Azevedo, C. S. y Antonini, Y. (2018). Does scale matter? The influence of three-level spatial scales on forest bird occurrence in a tropical landscape. PLoS One, 13(6), e0198732. https://doi.org/10.1371/journal.pone.0198732
Boelman, N. T., Asner, G. P., Hart, P. J. y Martin, R. E. (2007). Multi‐trophic invasion resistance in Hawaii: bioacoustics, field surveys, and airborne remote sensing. Ecological Applications, 17(8), 2137-2144. https://doi.org/10.1890/07-0004.1
Bradfer-Lawrence, T., Bunnefeld, N., Gardner, N., Willis, S. G. y Dent, D. H. (2020). Rapid assessment of avian species richness and abundance using acoustic indices. Ecological Indicators, 115, 106400. https://doi.org/10.1016/j.ecolind.2020.106400
Bradfer‐Lawrence, T., Gardner, N., Bunnefeld, L., Bunnefeld, N., Willis, S. G. y Dent, D. H. (2019). Guidelines for the use of acoustic indices in environmental research. Methods in Ecology and Evolution, 10(10), 1796-1807. https://doi.org/10.1111/2041-210X.13254
Breiman, L. (2001). Random forests. Machine learning, 45(1), 5-32. https://doi.org/10.1023/A:1010933404324
Brooks, M. E., Kristensen, K., van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., Skaug, H. J., Mächler, M. y Bolker, B. M. (2017). glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. https://doi.org/10.32614/RJ-2017-066
Carantón-Ayala, D. y Certuche-Cubillos, K. (2010). A new species of antpitta (Grallariidae: Grallaria) from the northern sector of the western Andes of Colombia. Ornitología Colombiana, 9, 56-70.
Carrara, E., Arroyo-Rodríguez, V., Vega-Rivera, J. H., Schondube, J. E., de Freitas, S. M. y Fahrig, L. (2015). Impact of landscape composition and configuration on forest specialist and generalist bird species in the fragmented Lacandona rainforest, Mexico. Biological Conservation, 184, 117-126. https://doi.org/10.1016/j.biocon.2015.01.014
Das, N., Mondal, A., Chaki, J., Padhy, N. y Dey, N. (2020). Machine Learning Models for Bird Species Recognition Based on Vocalization: A Succinct Review. Information Technology and Intelligent Transportation Systems, 117-124.
Depraetere, M., Pavoine, S., Jiguet, F., Gasc, A., Duvail, S. y Sueur, J. (2012). Monitoring animal diversity using acoustic indices: implementation in a temperate woodland. Ecological Indicators, 13(1), 46-54. https://doi.org/10.1016/j.ecolind.2011.05.006
Dröge, S., Martin, D. A., Andriafanomezantsoa, R., Burivalova, Z., Fulgence, T. R., Osen, K., ... y Kreft, H. (2021). Listening to a changing landscape: Acoustic indices reflect bird species richness and plot-scale vegetation structure across different land-use types in north-eastern Madagascar. Ecological Indicators, 120, 106929. https://doi.org/10.1016/j.ecolind.2020.106929
Elbers, J. (2011). Áreas protegidas de América Latina: situación actual y perspectivas para el futuro. Madrid, España: UICN-Unión Internacional para la Conservación de la Naturaleza y de los Recursos Naturales, 227p.
ESRI. (2016). ArcGIS Desktop: Release 10.5. Redlands, California, USA: Environmental Systems Research Institute
Ezzine de Blas, D., Rico, L., Ruiz Pérez, M. y Maris, V. (2011). La biodiversidad en el universo de los pagos por servicios ambientales: desentrañando lo inextricable. Revista Española de Estudios Agrosociales y Pesqueros, 228, 11-47.
Fahrig, L., Arroyo-Rodríguez, V., Bennett, J. R., Boucher-Lalonde, V., Cazetta, E., Currie, D. J. y Watling, J. I. (2019). Is habitat fragmentation bad for biodiversity? Biological Conservation, 230, 179-186. https://doi.org/10.1016/j.biocon.2018.12.026
Fahrig, L. (2003). Effects of habitat fragmentation on biodiversity. Annual review of ecology, evolution, and systematics, 34(1), 487-515.
Fairbrass, A. J., Rennert, P., Williams, C., Titheridge, H., & Jones, K. E. (2017). Biases of acoustic indices measuring biodiversity in urban areas. Ecological Indicators, 83, 169-177.
Garrigues, R. y Dean, R. (2014). The birds of Costa Rica, a field guide. Ithaca Nueva York, Cornell University Press.
Gonçalves, S., Pieretti, N., Young, R. J., & Duarte, M. (2021). Biophony in a noisy tropical urban forest fragment. Biota colombiana, 22(1), 96-107. https://doi.org/10.21068/c2021.v22n01a06
González-Maya, J. F., Víquez-R, L. R., Belant, J. L. y Ceballos, G. (2015). Effectiveness of Protected Areas for Representing Species and Populations of Terrestrial Mammals in Costa Rica. PLoS ONE, 10(5), e0124480. https://doi.org/10.1371/journal.pone.0124480
Hartig, F. (2022). DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models. R package version 0.4.5. https://CRAN.R-project.org/package=DHARMa
Hartigan, J. A., & Wong, M. A. (1979). Algorithm AS 136: A k-means clustering algorithm. Journal of the Royal Statistical Society. Series c (Applied statistics), 28(1), 100-108.
K. Lisa Yang Center for Conservation Bioacoustics at the Cornell Lab of Ornithology. (2022). Raven Pro: Interactive Sound Analysis Software (Version 1.6.3) [Computer software]. Ithaca, N. Y.: The Cornell Lab of Ornithology. https://ravensoundsoftware.com/
Kark, S. (2013). Effects of ecotones on biodiversity. Encyclopedia of biodiversity, 142-148. https://doi.org/10.1016/B978-0-12-384719-5.00234-3
Kasten, E. P., Gage, S. H. y Fox, J. (2012). The remote environmental assessment laboratory’s acoustic library: an archive for studying soundscape ecology. Ecological Informatics, 12, 50-67. https://doi.org/10.1016/j.ecoinf.2012.08.001
Keene, K. A. y Barlow, B. (2019). Benefits & Drawbacks to Variable-Radius Plots. Extension Alabama A&M & Auburn Universities. https://www.aces.edu/blog/topics/forestry/benefits-drawbacks-to-variable-radius-plots/
Kuhn, M. (2008). Building Predictive Models in R Using the caret Package. Journal of Statistical Software, 28(5), 1-26. https://doi.org/10.18637/jss.v028.i05
Metcalf, O. C., Barlow, J., Devenish, C., Marsden, S., Berenguer, E. y Lees, A. C. (2021). Acoustic indices perform better when applied at ecologically meaningful time and frequency scales. Methods in Ecology and Evolution, 12(3), 421-431. https://doi.org/10.1111/2041-210X.13521
Minello, M., Calado, L. y Xavier, F. C. (2021). Ecoacoustic indices in marine ecosystems: a review on recent developments, challenges, and future directions. ICES Journal of Marine Science, 78(9), 3066-3074. https://doi.org/10.1093/icesjms/fsab193
Mitchell, S. L., Bicknell, J. E., Edwards, D. P., Deere, N. J., Bernard, H., Davies, Z. G., & Struebig, M. J. (2020). Spatial replication and habitat context matters for assessments of tropical biodiversity using acoustic indices. Ecological Indicators, 119, 106717.
Nieto-Mora, D. A., Rodríguez-Buritica, S., Rodríguez-Marín, P., Martínez-Vargaz, J. D. y Isaza-Narváez, C. (2023). Systematic review of machine learning methods applied to ecoacoustics and soundscape monitoring. Heliyon, 9(10). https://doi.org/10.1016/j.heliyon.2023.e20275
Pérez‐Granados, C. y Traba, J. (2021). Estimating bird density using passive acoustic monitoring: a review of methods and suggestions for further research. Ibis, 163(3), 765-783. https://doi.org/10.1111/ibi.12944
Pieretti, N., Farina, A. y Morri, D. (2011). A new methodology to infer the singing activity of an avian community: the acoustic complexity index (ACI). Ecological Indicators, 11, 868-873. https://doi.org/10.1016/j.ecolind.2010.11.005
Porras, I. y Chacón-Cascante, A. (2018). Costa Rica’s Payments for Ecosystem Services Programme. International Institute for Environment and Development. https://www.iied.org/g04272
Pringle, R. M. (2017). Upgrading protected areas to conserve wild biodiversity. Nature, 546(7656), 91-99. https://doi.org/10.1038/nature22902
Rempel, R. S., Kaukinen, D. y Carr, A. P. (2012). Patch analyst and patch grid. Ontario Ministry of Natural Resources. Centre for Northern Forest Ecosystem Research, Thunder Bay, Ontario.
Renjifo, L. M. y Amaya-Villarreal, Á. M. (2017). Evolución del riesgo de extinción y estado actual de conservación de las aves de Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 41(161), 490-510. https://doi.org/10.18257/raccefyn.461
Rosli, Z., Zakaria, M. y Rajpar, M. N. (2018). Edge effects on foraging guilds of upperstory birds in an isolated tropical rainforest of Malaysia. JAPS: Journal of Animal & Plant Sciences, 28(1).
Sánchez-Chaves, O., & Navarrete-Chacón, G. (2017). La experiencia de Costa Rica en el pago por servicios ambientales: 20 años de lecciones aprendidas. Revista de Ciencias Ambientales, 51(2), 195-214. https://doi.org/10.15359/rca.51-2.11
Schulze, C. H. y Riedl, I. (2008). Bird assemblages of forested and human-modified countryside habitats in the Pacific lowlands of southern Costa Rica. Stapfia, 88(80), 395-408.
Slabbekoorn, H. (2004). Habitat-dependent ambient noise: consistent spectral profiles in two African forest types. The Journal of the Acoustical Society of America, 116(6), 3727-3733. https://doi.org/10.1121/1.1811121
Sueur, J., Aubin, T. y Simonis, C. (2008). Seewave, a free modular tool for sound analysis and synthesis. Bioacoustics, 18(2), 213-226. https://doi.org/10.1080/09524622.2008.9753600
Towsey, M., Wimmer, J., Williamson, I. y Roe, P. (2014). The use of acoustic indices to determine avian species richness in audio-recordings of the environment. Ecological Informatics, 21, 110-119. https://doi.org/10.1016/j.ecoinf.2013.11.007
Towsey, M. W. (2013). Noise removal from wave-forms and spectrograms derived from natural recordings of the environment. http://eprints.qut.edu.au/41131/
Valente, J. J. y Betts, M. G. (2019). Response to fragmentation by avian communities is mediated by species traits. Diversity and Distributions, 25(1), 48-60. https://doi.org/10.1111/ddi.12837
Vega-Hidalgo, A., Flatt, E., Whitworth, A. y Symes, L. (2021). Acoustic assessment of experimental reforestation in a Costa Rican rainforest. Ecological Indicators, 133, 108413. https://doi.org/10.1016/j.ecolind.2021.108413
Villanueva-Rivera, L. J. y Pijanowski, B. C. (2018). Soundecology: soundscape ecology. R package version. 1(3), 3. https://cran.r-project.org/web/packages/soundecology/soundecology.pdf
Villanueva-Rivera, L. J., Pijanowski, B. C., Doucette, J. y Pekin, B. (2011). A primer of acoustic analysis for landscape ecologists. Lanndscape Ecology, 26(9), 1233-1246. https://doi.org/10.1007/s10980-011-9636-9
Wildlife Acoustics Inc. (2023). Kaleidoscope User Guide. Maynard, Massachusetts. https://www.wildlifeacoustics.com/uploads/user-guides/Kaleidoscope-User-Guide.pdf
Yip, D. A., Bayne, E. M., Sólymos, P., Campbell, J. y Proppe, D. (2017). Sound attenuation in forest and roadside environments: Implications for avian point-count surveys. The Condor: Ornithological Applications, 119(1), 73-84. https://doi.org/10.1650/CONDOR-16-93.1
Anexo 1. Visualización de audio de ejemplo en el Sitio COR2 donde se muestra la presencia de sonidos de tipo “click” anormales.
Annex 1. Example audio display on the COR2 Site where the presence of abnormal “Click” type sounds is evident.

Nota: los sonidos aparecen en un alto porcentaje de las grabaciones, generalmente cuando hay una ligera lluvia de fondo, pero también cuando no la hay. Es probable que esta interferencia se deba a un deterioro de la cubierta protectora de los micrófonos, lo cual genera una exposición directa a la interferencia por contacto físico con los elementos del ambiente (Ej. ramas, gotas, hojas, viento, etc.).
1 Académico, Instituto Internacional en Conservación y Manejo de Vida Silvestre, Costa Rica. jimmybarrantesm@gmail.com; https://orcid.org/0000-0002-6088-638X
2 Académica, Instituto Internacional en Conservación y Manejo de Vida Silvestre, Costa Rica. mretamos@una.cr; https://orcid.org/0000-0002-4010-2998
3 Profesional Técnico, Departamento de Propuestas, Fondo Nacional de Financiamiento Forestal, Costa Rica. ricardo.bedoya@fonafifo.go.cr; https://orcid.org/0000-0002-9896-6895
4 Profesional Técnico, Departamento de Propuestas, Fondo Nacional de Financiamiento Forestal, Costa Rica. adriana.valerio@fonafifo.go.cr; https://orcid.org/0009-0009-5281-451X
5 Profesional Técnico, Departamento de Propuestas, Fondo Nacional de Financiamiento Forestal, Costa Rica. jonathan.sanchez@fonafifo.go.cr; https://orcid.org/0009-0001-8832-125X



Escuela de Ciencias Ambientales,
Universidad Nacional, Campus Omar Dengo
Apartado postal: 86-3000. Heredia, Costa Rica
Teléfono: +506 2277-3688
Correo electrónico revista.ambientales@una.ac.cr