Rev. Mar. y Cost. ISSN 1659-455X. Vol. 1. 111-125, Diciembre 2009.
DOI: https://doi.org/10.15359/revmar.1.6
EVALUACIÓN DE LAS ASOCIACIONES DE PECES DE LOS ARRECIFES CORALINOS DEL CENTRO INTERNACIONAL DE BUCEO MARÍA LA GORDA, PARQUE NACIONAL GUANAHACABIBES, CUBA
Dorka Cobián Rojas 1* y Pedro Pablo Chevalier Monteagudo 2
1 Parque Nacional Guanahacabibes, Centro de Investigaciones y Servicios Ambientales (ECOVIDA), Ministerio de Ciencia, Tecnología y Medio Ambiente (CITMA). La Bajada, Código Postal 22 100, Pinar del Río, Cuba. email: dorkacobian@gmail.com*
2 Acuario Nacional de Cuba, Ministerio de Ciencia, Tecnología y Medio Ambiente (CITMA). 1ra. y 60, Miramar, Playa, Ciudad de La Habana, Código Postal 12 100, Cuba.
Recibido 16-IV-2009
Aceptado 28-IX-2009
RESUMEN
El buceo contemplativo en los arrecifes coralinos es una de las pocas actividades autorizadas en el área manejada por el Centro Internacional de Buceo María la Gorda del Parque Nacional Guanahacabibes. Se evaluó la composición y estructura de las asociaciones de peces en los arrecifes utilizados por el Centro Internacional de Buceo María la Gorda. Considerando la posibilidad de cambios estacionales en la densidad y composición de los peces, se realizaron muestreos en las épocas seca y lluviosa. Se realizaron censos visuales en ocho sitios de buceo (12 transectos de 50 x 2 m en cada uno), en dos biotopos característicos del arrecife. Se estimaron los índices de diversidad, equitatividad, riqueza de especies, densidad y biomasa de peces. Los valores de densidad promedio y los índices de diversidad por estaciones no mostraron diferencias significativas entre las épocas. Se registraron 134 especies en toda el área. La abundancia de peces fue superior en los sitios de mayor complejidad estructural, correspondientes a la zona de camellones de la pendiente arrecifal. Se observó un predominio de peces de pequeña talla y pobre presencia de peces de mediana y gran talla. Las familias con mayor densidad fueron Pomacentridae y Labridae, y las de mayor biomasa Lutjanidae y Labridae. La estructura trófica estuvo dominada por los plantófagos y omnívoros. Aunque se observaron carnívoros de tallas medianas, su densidad fue inferior al resto de los grupos, lo cual parece ser consecuencia de la pesca furtiva.
Palabras claves: Asociaciones de peces, diversidad, arrecifes coralinos, Cuba.
ABSTRACT
Recreational SCUBA diving on the coral reefs is one of the few activities allowed in the area managed by the Centro Internacional de Buceo María la Gorda at the Guanahacabibes National Park. The composition and structure of fish associations were evaluated on the reefs visited by users of the diving center. To assess possible seasonal changes in fish community densities and composition surveys were carried during the dry and rainy seasons. Visual censuses were carried out at eight diving sites comprising two characteristic reef biotopes. Indices for diversity, evenness, species richness, density, and fish biomass were estimated. Mean density values by sampling site did not show significant differences between seasons. A total of 134 species were recorded in the area. Fish abundances were greater in sites of higher structural complexity, matching areas of ridges on the reef slope. Prevalence of small-sized fish and a general poor presence of medium to high-sized fish were observed. Fish families with the higher densities were Pomacentridae and Labridae, and those with the higher biomass were Lutjanidae and Labridae. The trophic structure was dominated by phytophagous and omnivorous fish. Medium-sized carnivorous fish were observed, however, their densities were much lower than for the rest of the trophic groups, which could be a consequence of illegal fishing activities.
Keywords: Fish associations, diversity, coral reefs, Cuba.
INTRODUCCIÓN
El área de arrecifes coralinos del Centro Internacional de Buceo María la Gorda, en el Parque Nacional Guanahacabibes, fue designada por el Ministerio de la Industria Pesquera como Zona Bajo Régimen Especial de Uso y Protección desde el 2003, en la cual solo se permite la actividad de buceo contemplativo y pesca de subsistencia únicamente en los sectores designados para estas actividades. Sin embargo, no existe aún un estudio detallado de la biodiversidad y los factores que la afectan, los que constituyen elementos fundamentales para la implementación y control del Plan de Manejo existente.
Los arrecifes del área han sido considerados entre los de mayor belleza y mejor estado de conservación del país (Alcolado et al. 2003; Gotera, 2005), y a juzgar por el monitoreo que se efectúa regularmente (datos no publicados) se mantienen en igual condición. Sin embargo, algunas evidencias indican que la ictiofauna de esos arrecifes puede estar afectada por la pesca ilegal (Claro y Cantelar, 2003), aunque se ha reducido después de la creación del Parque Nacional y el incremento de la vigilancia. Durante el período de mayo, junio y julio se autoriza en el área la pesca artesanal sobre las agregaciones de desove del pargo criollo, Lutjanus analis, por los escasos residentes de las comunidades aledañas.
En julio de 1999 Claro y Cantelar (2003) realizaron la primera evaluación de las asociaciones de peces en cuatro arrecifes someros (a 5-8 m y 9-13 m de profundidad) utilizando la metodología AGRRA (solo especies seleccionadas; AGRRA, 1999) y en tres de esos arrecifes, mediante censos de todas las especies (Brock, 1954). Con ambos métodos encontraron valores relativamente bajos de densidad y biomasa. No obstante, no se han realizado evaluaciones en arrecifes más profundos, donde se desarrolla la principal actividad de buceo contemplativo. Tampoco existe información en cuanto a la composición de la ictiofauna en relación con diferentes características del arrecife, en particular con su complejidad estructural.
Además, algunos autores argumentan la necesidad de evaluar la estructura de la comunidad en diferentes épocas del año, considerando que esta puede variar estacionalmente (Aguilar y González-Sansón, 1998; Aguilar et al. 2004; Hernández et al. 2006). La aclaración de este aspecto resulta de gran importancia metodológica, por cuanto su indefinición provoca dudas en cuanto a la variabilidad de los resultados de muestreos realizados en diferentes momentos del ciclo anual.
El presente trabajo tuvo como objetivos validar la hipótesis de la existencia de diferencias estacionales en la diversidad y densidad de peces en los arrecifes coralinos durante los dos principales períodos climáticos en Cuba y aplicar los resultados a la evaluación de la estructura de dicha comunidad en los arrecifes del sector del área marina protegida utilizada por el turismo, como elemento básico para su manejo y conservación. Los resultados obtenidos fueron incorporados al Plan de Manejo en vigor para el área y se mantiene el monitoreo de esta.
MATERIALES Y MÉTODOS
El Centro Internacional de Buceo María la Gorda se encuentra ubicado en el área marina del Parque Nacional Guanahacabibes, con una extensión de 15.950 ha que abarca la porción sur de la Reserva de Biosfera, desde Jaimanitas al este de Cabo Corrientes hasta Cabo San Antonio.
En dos meses representativos de los períodos de sequía-invierno (diciembre del 2003) y lluvias-verano (septiembre del 2004), se realizaron censos visuales mediante buceo autónomo en ocho de los puntos de buceo más visitados por los buzos (Figura 1).
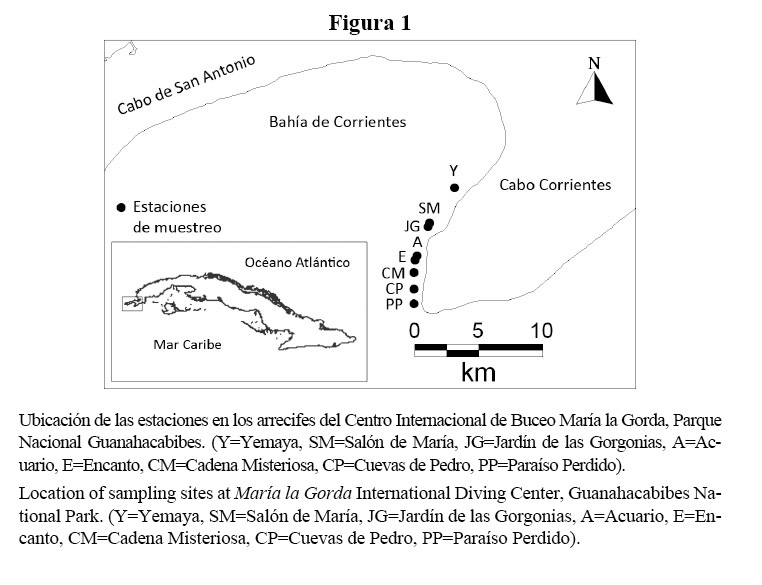
En esta zona, los arrecifes son de tipo franjeantes o costeros y su perfil característico generalmente es el de una terraza única (González-Ferrer et al. 2007) que culmina en un escarpe profundo de estructura variada. Se seleccionaron dos subzonas del arrecife: 1. cabezos y parches coralinos, ubicados en una terraza rocosa con parches de arena que forman una franja paralela a la línea de costa, a profundidades entre 6 y 10 m; 2. camellones y canales que se localizan a profundidades entre 15 y 20 m (Cuadro 1).
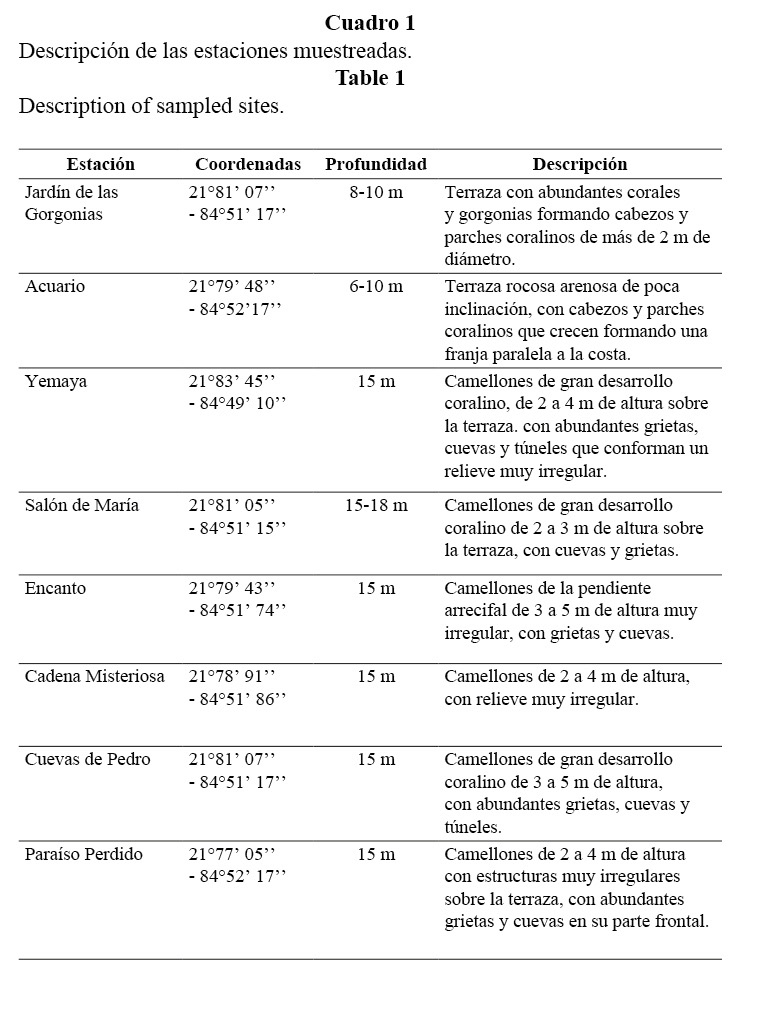
En cada estación se realizaron seis transectos de 50 m de largo por 2 de ancho, anotándose todas las especies presentes (Brock, 1954), cubriendo un área total de 600 m2. Se registraron el número de individuos de cada especie y su talla estimada por personal previamente entrenado. Con esos valores se calculó el peso de los individuos, empleando para ello las relaciones largo-peso obtenidas en la literatura (Bohnsack y Harper, 1988; Claro y García-Arteaga, 1994; García-Arteaga et al. 1997), para hacer estimados de la biomasa. Los datos se procesaron por el programa ESTRUCOM (Instituto de Oceanología), obteniéndose el listado en orden taxonómico de las especies observadas, densidad y biomasa por especie y familia, el índice de diversidad H’ (Shannon y Weaver, 1949), el índice de riqueza R1 (Margalef, 1951) y el índice de equitatividad J’ (Pielou, 1966). Para cada estación y zona del arrecife se obtuvo el valor promedio de densidad (individuos/m2) y biomasa (g/m2) de peces en los seis transectos ± el error estándar (EE).
La estructura de la comunidad se analizó además mediante el método de frecuencia-abundancia (F-A) descrito por Mason y Bryant (1974) para combinar en un solo indicador, la abundancia en número de cada especie, con su frecuencia de aparición en los muestreos. Para ello se establecieron arbitrariamente tres niveles de frecuencias: A. especies que aparecen en más del 67% de los muestreos; B. entre 33% y 67% y C. en menos del 33%. El porcentaje en densidad se dividió a su vez en cuatro niveles: 1. más del 10%; 2. entre 1 y 10%; 3. entre 1.1 y 1%; 4. menos de 0.1%. De esta forma, cada especie se coloca en una de las doce categorías o combinaciones de frecuencia-abundancia-biomasa de acuerdo con su presencia y con su densidad.
Adicionalmente se estimó el índice de jerarquía IJ (Ranking Index) para cada especie, multiplicando el porcentaje de frecuencia por el porcentaje de individuos y se jerarquizan después los valores obtenidos.
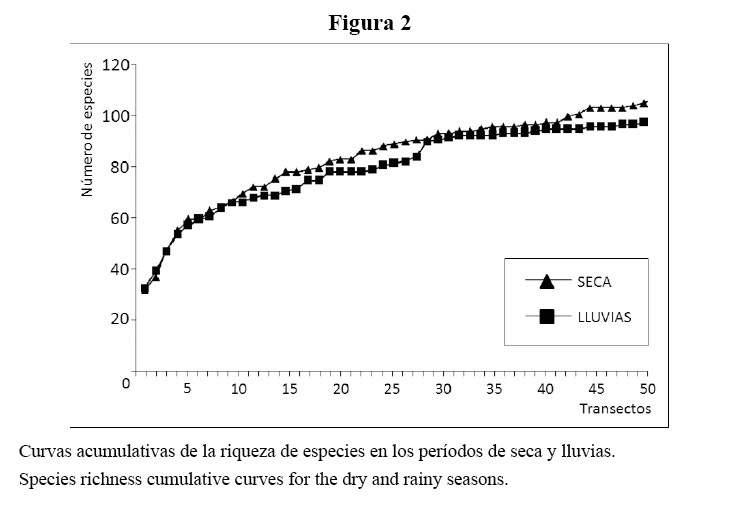
Se construyeron curvas acumulativas de riqueza de especies para cada época del año, mediante combinaciones aleatorias del número acumulado de especies contra el número de transectos muestreados en toda el área en cada etapa.
Los datos fueron comparados mediante estadística descriptiva (medias, error estándar). Los valores de densidad promedio fueron procesados mediante análisis de varianza (ANOVA) considerando un diseño de clasificación simple. Para la comparación múltiple a posteriori de las medias de las densidades, se utilizó la prueba de Newman-Keuls. Se verificaron las asunciones de homogeneidad de varianza y normalidad de los datos, siguiendo los criterios de Zar (1996) y Underwood (1997). Se utilizó siempre un índice de significación de 0.05 con intervalos de confianza al 95% y la transformación utilizada resultó T = 10.48/2 = 0.76. Los valores de densidad de las dos zonas muestreadas fueron comparados mediante la prueba t Student. Todos los datos fueron procesados mediante el programa de STATISTICA 6.0 (StatSoft, 2001).
Se adoptaron las categorías tróficas propuestas para cada especie por Sierra et al. (1994), con base en numerosos estudios sobre la alimentación de los peces del Caribe, que considera como criterio el consumo, en más de 70% de determinado componente y en casos excepcionales 50% para los que utilizan dos tipos de alimento principal, cuya proporción cambia con el crecimiento, como es el caso de los ictiobentófagos.
RESULTADOS
Las curvas de acumulación de riqueza de especies en los períodos de seca y lluvias alcanzaron un nivel cercano al asintótico, lo que evidencia, además de su similitud, que el número de transectos muestreados ofrece un estimado confiable de la riqueza de especies (Figura 2).
Se encontraron 134 especies de peces, aunque el número en cada estación fluctuó entre 45 y 69 (56 ± 4.1 EE). Los arrecifes de cabezos y parches (Acuario y Jardín de las Gorgonias) presentaron valores más elevados de riqueza de especies y del índice de equitatividad. En los camellones, donde la complejidad estructural del sustrato es mayor, se encontraron valores más bajos de H’ y J’, y densidades más altas (Cuadro 2).
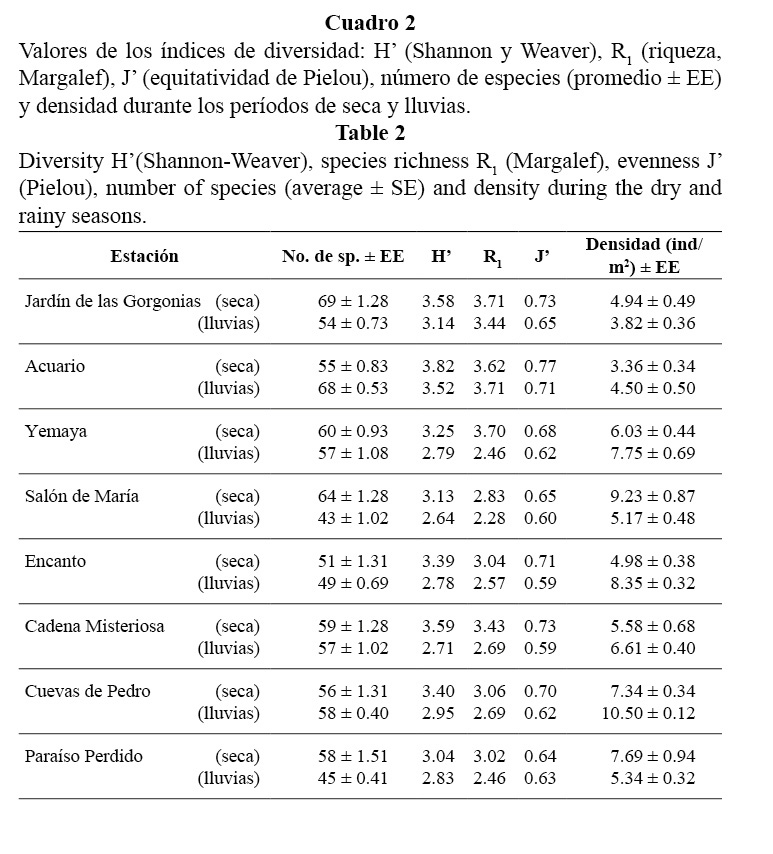
No se observaron diferencias significativas entre los períodos de seca y lluvias (Cuadro 2) para los valores de densidad (F = 0.3536; p = 0.5509). Las diferencias observadas en algunas estaciones no son regulares ni guardan una clara relación con la influencia de las lluvias. Por lo tanto, se unieron los datos obtenidos en ambos períodos, para obtener una muestra más representativa de la estructura de la comunidad de peces.
Al comparar la densidad por estaciones, la prueba de Newman-Keuls solo mostró diferencias significativas (F = 2.2806; p = 0.0359) en dos estaciones (Figura 3).
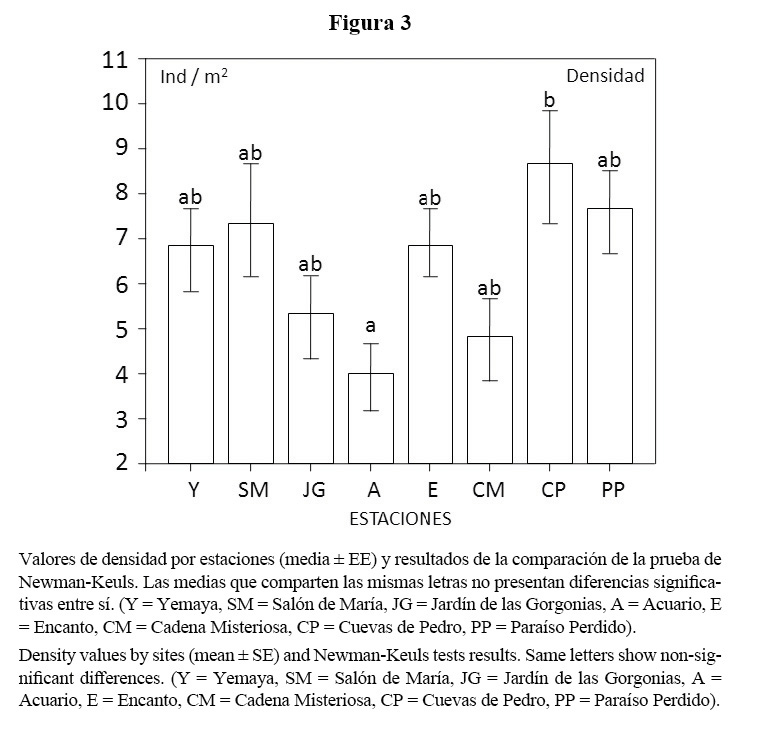
Los valores de densidad obtenidos entre biotopos (cabezos y parches coralinos y camellones) presentaron diferencias significativas (T = 2.4693; p = 0.0153). Los últimos presentaron mayores valores de densidad (7.16 ± 0.84 ind/m2) que los cabezos y parches coralinos (4.15 ± 0.82 ind/m2).
Los valores promedios de la densidad para las especies que representaron el 95% del total de los individuos en toda la zona estudiada fueron ordenados de forma decreciente, según el índice jerarquía (Cuadro 3).
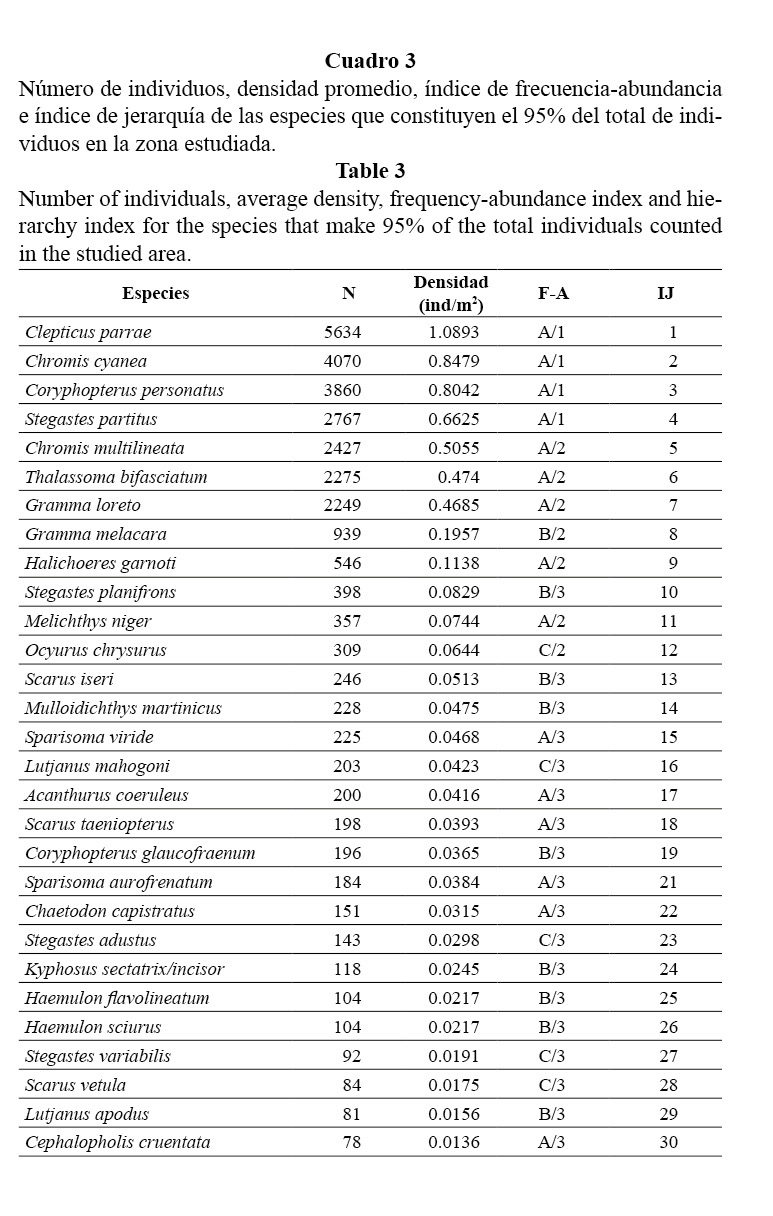
En ambas zonas del arrecife, las familias con mayor densidad fueron Pomacentridae y Labridae. En los parches hubo una abundancia mayor de roncos por la presencia de Haemulon sciurus y Haemulon flavolineatum. La densidad de meros y pargos fue baja (Figura 4).
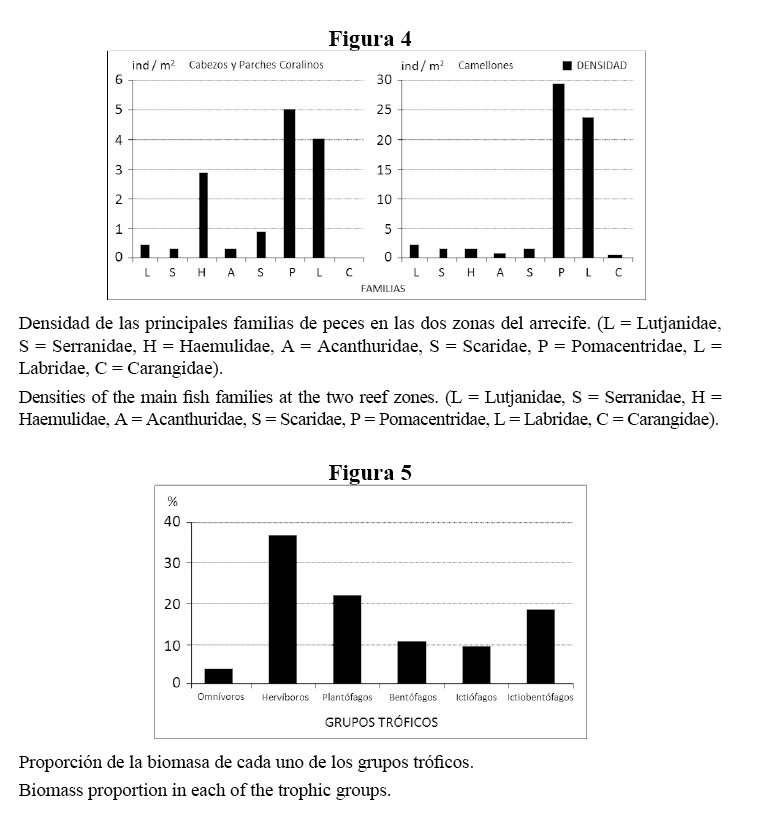
De acuerdo con la proporción de biomasa por categorías tróficas de los peces observados en toda la zona (Figura 5), los herbívoros (donde predominan Sparisoma viride, Scarus vetula, Acanthurus coeruleus y Kyphosus sectatrix/incisor) y los plantófagos (Clepticus parrae, Chromis cyanea, Chromis multilineata y Gramma loreto) presentaron biomasas más altas, producto de un elevado número de individuos de estas especies. Le siguen los ictiobentófagos, por el aporte de Ocyurus chrysurus de tallas relativamente grandes entre 25 y 35 cm de longitud, que forman agregaciones en el borde de la terraza profunda. También fue considerable la presencia de Lutjanus apodus y Cephalopholis cruentata.
DISCUSIÓN
Los valores de densidad de peces en los períodos de seca y lluvias no presentaron diferencias significativas, salvo en dos de las estaciones, pero tales diferencias no guardan una relación definida con la estación del año, ya que en unos casos fue mayor en lluvias y en otros en seca. Por lo tanto, se rechaza la hipótesis de variaciones estacionales en la densidad de peces en el área estudiada. Las diferencias observadas parecen ser consecuencia de los movimientos y los cambios en la distribución de los peces en el arrecife.
La composición por especies de la zona presentó características similares a la encontrada en otros arrecifes de Cuba y el Caribe (Claro et al. 1998; Chevalier y Cárdenas, 2005; Hernández et al. 2006). Las especies más abundantes fueron: Chromis cyanea, Stegastes partitus, Clepticus parrae, Thalassoma bifasciatum y Coryphopterus personatus/hyalinus, casi todas de pequeña talla (menores de 12 cm LT), pero de gran colorido y atractivo para el buceo contemplativo.
La existencia de valores más bajos de H’ y J’ en los camellones y cangilones parece ser consecuencia de una alta dominancia de Coryphopterus personatus/hyalinus, Gramma loreto, Chromis cyanea, Chromis multilineata, Stegastes partitus, Clepticus parrae y Thalassoma bifasciatum, que conformaron el 73% de todos los individuos.
La diversidad en las comunidades de peces puede estar dada por la complejidad del biotopo (refugios), la cercanía del arrecife principal (fuente de inmigración), las diferencias del subnicho alimentario, las respuestas adaptativas de competición y depredación, entre otras (Alcolado, 1984). Los cabezos y parches coralinos presentaron mayores valores de los índices de diversidad y equitatividad que los camellones, lo cual parece ser consecuencia de una alta dominancia de unas pocas especies en los camellones, donde además, el fondo presentó mayor diversidad estructural. Resultados similares fueron encontrados por Chevalier y Cárdenas (2005) en la Bahía de Cochinos, quienes plantearon que los índices de diversidad y equitatividad disminuyen con la profundidad y el aumento de la complejidad del sustrato.
Los valores medios de densidad en los parches (4.15 ± 0.82 ind/m2) fueron similares a los obtenidos por Claro y Cantelar (2003) en 1999 (5.41 ± 0.22 ind/m2) en tres estaciones. No obstante, la densidad de herbívoros (1.05 ± 0.02 ind/m2) y de pargos y meros (ictiobentófagos) fue mayor en nuestros muestreos (0.62 ± 0.01 ind/m2) que la reportada por dichos autores (0.36 ± 0.02 ind/m2), lo cual podría indicar una cierta recuperación en la abundancia de peces entre uno y otro período.
Las estaciones de Paraíso Perdido y Cuevas de Pedro presentaron valores elevados de densidad y mayores proporciones de biomasa por grupos tróficos de esta misma región. Estos dos sitios poseen mayor desarrollo coralino que los demás estudiados con similar sustrato, los cuales brindan refugio a meros y pargos de mayor talla. Además, estas estaciones se encuentran más alejadas de la costa y menos accesibles a los pescadores furtivos de las comunidades aledañas.
Con respecto a la biomasa, el gremio dominante fue el de los herbívoros, destacándose Sparisoma viride, Scarus vetula, Acanthurus coeruleus y Kyphosus sectatrix/incisor. Esta última especie tuvo una alta incidencia en este indicador, ya que se observaron numerosos individuos de tallas entre 25 y 35 cm de longitud, consideradas como altas en comparación con las observadas en otros herbívoros.
Se observaron carnívoros de tallas relativamente grandes (30-50 cm), principalmente Lutjanidae y Serranidae, en el área de los camellones, sin embargo, su densidad fue baja. Esto parece ser consecuencia de la pesca submarina ilegal que se realiza en la zona, aunque debe destacarse que las regulaciones establecidas por el Parque Nacional han reducido estas actividades. Sin embargo, los valores de densidad y biomasa son elevados si se comparan con el norte de la Ciudad de La Habana, la cual está sobrepescada y dominada por especies pequeñas, de nivel trófico bajo, con ausencia casi total de estas familias (Aguilar, 2005; Durán y Claro, en prensa).
La relativamente alta abundancia de peces de pequeña (5-15 cm) y mediana talla (15-30 cm), pero de gran atractivo por su colorido y conducta, como los parches, angelotes, isabelitas, cromis y loretos, es un factor importante en el valor estético del área para las actividades de buceo contemplativo.
Caballero et al. (2007) y González-Ferrer et al. (2007) argumentaron que no han existido afectaciones en la diversidad y densidad de corales por las actividades de buceo en el área. Durante nuestros muestreos, realizados en las mismas estaciones analizadas por estos autores, no se observaron señales de conducta huidiza o reacciones negativas por parte de los peces ante la presencia de los buceadores, aunque este factor debe ser evaluado cuantitativamente. En la actualidad, el número de visitantes no excede la capacidad de carga aceptable para actividades de buceo en parques nacionales (4.000 buzos por punto de buceo por año, según Hawkins y Roberts, 1997). Sin embargo, es muy probable que en el futuro aumente el número de buceadores, por lo que es importante el monitoreo de la zona de buceo, para evaluar posibles cambios en la biodiversidad y conducta de los peces.
AGRADECIMIENTOS
Agradecemos a los biólogos Dámaso Rosales, Daylin Muñoz, Karel Cantelar y Antonio Cárdenas, por su colaboración en la toma de datos, y a Rodolfo Claro Madruga del Instituto de Oceanología por sus útiles señalamientos y comentarios.
BIBLIOGRAFÍA
AGRRA. (1999). Atlantic and Gulf Rapid Reef Assessment (AGRRA). Mesoamerican Reef. System Workshop, May 17-21. 37 pp.
Aguilar, B. C. (2005). La ictiofauna de la Ciudad de La Habana: Efectos acumulativos de agentes estresantes múltiples en varios niveles de organización biológica. Tesis de Doctorado. Centro de Investigaciones Marinas, Universidad de La Habana.
Aguilar, C. y González-Sansón, G. (1998). Variación estacional de la abundancia de juveniles de peces en una zona litoral del sublitoral rocoso de La Habana, Cuba. Rev. Invest. Mar. 19(1), 38-45.
Aguilar, C., González-Sansón, G., Munkittrick, K. R. and Maclatchy, D. L. (2004). Fish assemblages on fringe coral reefs of the northern coast of Cuba near Havana. Harbor. Ecotox. Env. Saf., 58, 126-138.
Alcolado, M. P., Martínez-Daranas, B., Menéndez-Macía, G., del Valle, R., Hernández, M. and García, T. (2003). Rapid assessment of coral communities of María la Gorda, southeast Ensenada de Corrientes, Cuba (part 1: stony corals and algae). Pp. 268-277. En Status of Coral Reefs in the Western Atlantic: Results of Initial Surveys, Atlantic and Gulf Rapid Reef Assessment (AGRRA), Judith C. Lang (ed.). Program. Atoll. Res. Bull. 496, 630 pp.
Alcolado, P. M. (1984). Utilidad de algunos índices ecológicos estructurales en el estudio de comunidades marinas de Cuba. Cien. Biol., 11, 61-77.
Bohnsack, J. A. and Harper, D. E. (1988). Length-weight relationships of selected marine reef fishes from the southeastern United States and the Caribbean. NOAA Tech. Mem.
Brock, V. E. (1954). A preliminary report on a method of estimating reef fish populations. J. Wildlife Manage., 18, 297-308.
Caballero, H., González-Ferrer, S., Cobián, D., Álvarez, S. y Alcolado, P. M. (2007). Evaluación AGRRA en diez sitios de buceo de “María la Gorda”, Bahía Corrientes, Cuba. Rev. Invest. Mar. 28(2), 131-138.
Chevalier, P. y Cárdenas, A. (2005). Variación espacial y temporal de las asociaciones de peces en arrecifes coralinos de la costa de la Bahía de Cochinos. I: Abundancia y diversidad. Rev. Invest. Mar. 26(1), 45-57.
Claro, R. y García-Arteaga, J. P. (1994). Estructura de las comunidades de peces en los arrecifes del grupo insular Sabana-Camagüey, Cuba. Avicennia, 2, 83-107.
Claro, R. and Cantelar, K. (2003). Rapid assessment of coral communities of María la Gorda, southeastern Ensenada de Corrientes, Cuba (part 2: reef fishes). Atoll. Res. Bull., 496, 278-293.
Claro, R., García-Arteaga, J. P., Bouchon, Y., Louis, M. y Bouchon C. (1998). Caracterización de la estructura de las comunidades de peces en los arrecifes de las Antillas Menores y Cuba. Avicennia, 8/9, 68-86.
Durán, A. y Claro, R. (en prensa). Actividad alimentaria de los peces herbívoros y su impacto en arrecifes con diferente nivel de degradación antrópica. Rev. Biol. Trop.
García-Arteaga, J. P., Claro, R. and Valle, S. (1997). Length-weight relationships of Cuban Marine fishes. NAGA, The ICLARM Quartely, 20(1), 38-42.
González-Ferrer, S., Caballero, H., Alcolado, P. M., Jiménez, A., Martín, F. y Cobián, D. (2007). Diversidad de corales pétreos en once sitios de buceo recreativo de “María la Gorda”, Cuba. Rev. Invest. Mar. 28(2), 121-130.
González-Sansón, G., Aguilar, C., Angulo, J. y González, C. (1997) a. Variación espacial y estacional de la ictiofauna en un arrecife de coral costero de la región noroccidental de Cuba. II: Diversidad. Rev. Invest. Mar., 18(3), 233-240.
Gotera, G. G. (2005). Buceando en Cuba/ Diving in Cuba. Ediciones Niocia, S. L. Catalunya. Barcelona, España. 146 pp.
Hawkins, J. P. and Roberts, C. M. (1997). Estimating the carrying capacity of coral reefs for recreational scuba diving. Proc. 8th Int. Coral Reef Symp., 2, 1923-1926.
Hernández, I., Consuelo, A. y González-Sansón, G. (2006). Variación estacional de la abundancia de especies de peces seleccionadas en el sublitoral rocoso de Ciudad de La Habana, Cuba. Rev. Invest. Mar., 27(1), 61-68.
Margalef, R. (1951). Diversidad de especies en las comunidades naturales. Publ. Inst. Biol. Apl. Barcelona, 9, 5-27.
Mason, C. F. and Bryant, R. J. (1974). The structure and diversity of the animal communities in a broadland reeds-swamp. J. Zool., 172, 289-302.
Pielou, E. C. (1966). The measurement of diversity in different types of biological collections. J. Theoret. Biol. 13, 13-144.
Shannon, C. E. and Weaver, W. (1949). The mathematical theory of communication. University of Illinois Press. 117 pp.
Sierra, L. M., Claro, R. y Popova, O. A. (1994). Alimentación y relaciones tróficas. En: R. Claro (ed.), Ecología de los peces marinos de Cuba, pp. 263-320. Instituto de Oceanología, Academia de Ciencias de Cuba y Centro de Investigaciones de Quintana Roo (CIQRO), México.
StatSoft, Inc. (2001). STATISTICA (data analysis software system), version 6. www.statsoft.com.
Underwood, A. J. (1997). Experiments in ecology. Their logical design and interpretation using analysis of variance. Cambridge University Press, Cambridge. 504 pp.
Zar, J. H. (1996). Biostatistical analysis. 3ra. Ed. Prentice Hall, New Jersey. 662 pp.

Revista Ciencias Marinas y Costeras está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional.