ISSN: 1659-455X • e-ISSN: 1659-407X
Vol. 16 (1), enero-junio 2024
Recepción 15 enero 2024 • Corregido 20 mayo 2024 • Aceptado 23 mayo 2024
DOI: https://dx.doi.org/10.15359/revmar.16-1.1
 |
Estructura comunitaria de peces, asteroideos y equinoideos en el Farallón de San Ignacio, arrecife situado en la “brecha faunística de Sinaloa”, México Community structure of asteroid and echinoid fish at the Farallón de San Ignacio reef located at the “Sinaloan gap”, Mexico Héctor Reyes-Bonilla1, Violeta Martínez-Castillo1,2*, Bárbara Rojas-Montiel1,4 y Manuel Olán-González1,3 |
RESUMEN
Aunque la investigación sobre los arrecifes rocosos del golfo de California ha sido extensa, existen sitios sobre los cuales se tiene escasa información como el Farallón de San Ignacio, reconocido por su alta biodiversidad y por localizarse en la zona de transición biogeográfica llamada “brecha faunística de Sinaloa”. Debido a pocos datos disponibles de este punto, el objetivo del presente trabajo fue describir la composición y estructura de los ensamblajes de peces, asteroideos y equinoideos del Farallón de San Ignacio, para lo cual se realizaron censos visuales en noviembre de 2016, mediante transectos de banda de 25 m de largo por 4 m de ancho, para peces, y 25 m de largo por 2 m de ancho, para invertebrados. Se calcularon índices de abundancia, riqueza, diversidad y uniformidad, así como se compararon con comunidades insulares situadas a la misma latitud, en la costa oeste del golfo de California, visitadas el mismo año. Se registraron 34 especies de peces, dos de asteroideos y cuatro de equinoideos. Las especies más abundantes fueron el pez Azurina atrilobata, la estrella Phataria unifascialis y el erizo Diadema mexicanum. A pesar de su aislamiento, la complejidad de la estructura comunitaria del Farallón es similar a la de otras islas en el golfo de California, aunque hubo discrepancias en la composición de especies, debido a la alta incidencia de herbívoros en este lugar.
Palabras clave: abundancia, diversidad, golfo de California, herbivorismo, riqueza.
ABSTRACT
Even though there is extensive research on the rocky reefs in the Gulf of California, some areas still have scarce information, such as the Farallón de San Ignacio, which is well known for its high biodiversity and its location in the biogeographic transition zone known as the “Sinaloa gap.” Due to the limited information available for this site, the present study aimed to describe the composition and structure of asteroid and echinoid fish assemblages at the Farallón de San Ignacio. For this purpose, visual surveys were conducted in November 2016 using belt transects of 25 m long by 4 m wide for fish and 25 m long by 2 m wide for invertebrates, to estimate species abundance, richness, diversity, and evenness indices, and to compare them with other rocky reefs located at the same latitude on the western coast of the Gulf of California that were visited the same year. A total of 34 species were recorded, including two asteroids and four echinoids. The most abundant species were the fish Azurina atrilobata, the starfish Phataria unifacialis, and the sea urchin Diadema mexicanum. Despite the Farallón’s isolation, the complexity of its community structure is similar to that of other islands in the Gulf of California, although there were differences in species composition due to the high abundance of herbivorous species at this site.
Keywords: abundance, diversity, Gulf of California, herbivores, species richness.
En las últimas dos décadas, el conocimiento sobre la estructura comunitaria de la ictiofauna y de diversos grupos de invertebrados presentes en arrecifes rocosos y coralinos del golfo de California ha aumentado considerablemente. Se tienen numerosos trabajos que presentan información sobre aspectos como riqueza, abundancia y diversidad ecológica para localidades de la costa oeste de este cuerpo de agua, especialmente, entre las latitudes 25° N y 22° N, en el caso de los peces (Pérez-España et al. 1996; Aburto-Oropeza & Balart, 2001; Álvarez-Filip et al. 2006; Barjau-González et al. 2012); en áreas más extensas, en el de los invertebrados, como asteroideos y equinoideos, y, recientemente, también en el de los ofiuroideos 29° N a 24° N (Holguín-Quiñones et al. 2000; González-Medina et al. 2006; Herrero-Pérezrul et al. 2007; Holguín-Quiñones et al. 2008; Luna-Salguero & Reyes-Bonilla, 2010; Granja-Fernández et al. 2023; Maya-Alvarado et al. 2023). Además, existen análisis comparativos a escalas más amplias, que abarcan prácticamente la totalidad del golfo y contrastan la estructura comunitaria de los grupos taxonómicos citados (Reyes-Bonilla et al. 2005; Herrero-Pérezrul et al. 2009; Ulate et al. 2016; Granja-Fernández et al. 2023; Maya-Alvarado et al. 2023), aunque se centran, más que todo, en la costa occidental, caracterizada por la presencia de arrecifes rocosos y coralinos (Reyes-Bonilla & López-Pérez, 2009). No obstante, hay aún zonas prácticamente desconocidas, desde la óptica del estudio de zonas arrecifales, de modo particular, en los litorales de los estados de Sonora y Sinaloa (costa oriental del golfo de California).
En gran medida, la ausencia de información para las áreas citadas es una consecuencia lógica de la fisiografía general de esa región, dominada por largas extensiones de costa arenosa, manglares y lagunas costeras, que impiden el establecimiento de especies asociadas a fondos duros; el cambio en la composición del fondo con respecto a zonas más sureñas como bahía de Banderas (20°N) o norteñas como Guaymas (27°N), ambas con fondo rocoso predominante, es tan abrupto que constituye un filtro biogeográfico, el cual ha sido llamado “brecha faunística de Sinaloa” (“Sinaloan gap”, según Hastings, 2000; Robertson & Cramer, 2009). A pesar de ello, existen pequeños parches de fondo firme en islas cercanas a las ciudades de Mazatlán (23°N) y Topolobampo (25°N); los de la primera área han sido descritos de forma somera en algunas publicaciones (Caso-Muñóz et al. 1996; Cupul-Magaña, 2003; Medina-Rosas, 2006), mientras que para el segundo sitio las formaciones de fondo duro no han sido caracterizadas y solo se da la mención de la presencia de los arrecifes en zonas adyacentes, en específico, en la isla Farallón de San Ignacio (referida en el texto como “el Farallón”). Según Brusca (1980), este espacio albergaba una de las comunidades biológicas más diversas en el golfo de California, pero, por su relativo aislamiento (la isla se sitúa aproximadamente a 30 km al oeste de la costa más cercana), los ensamblajes de peces e invertebrados permanecen aún sin describir. De este punto, solo hay datos sobre la presencia de poblaciones de lobo marino Zalophus californianus (Lesson, 1828) y diversas aves marinas (González-Bernal et al. 2002; Guevara-Medina et al. 2008).
El presente estudio representa un avance en el conocimiento de una zona relevante, al representar una frontera biogeográfica poco explorada del golfo de California, y su principal objetivo fue describir tanto la composición de la fauna como la estructura comunitaria de tres grupos de organismos marinos presentes en el Farallón de San Ignacio: peces óseos, asteroideos y equinoideos. Como complemento, estos datos fueron contrastados cualitativa y cuantitativamente con los registrados en arrecifes rocosos que se ubican en latitudes similares localizadas en la costa oeste del golfo de California.
Los datos base del presente estudio se generaron en una visita al Farallón de San Ignacio (25° 25’ N, 109° 22’ O, Fig. 1) el 15 de noviembre de 2016, como parte de una expedición que recorrió arrecifes rocosos costeros e insulares del golfo de California, desde isla Cerralvo, en el sur, hasta isla Ángel de la Guarda, en el norte. El Farallón de San Ignacio es un promontorio rocoso que se encuentra en la parte suroriental del golfo de California, a 36 km de la costa de Topolobampo, Sinaloa y cerca de la cuenca de Farallón. La isla es muy pequeña y se rodea de aguas profundas; abarca una superficie de solo 16 ha (Samaniego-Herrera et al. 2009), con un perímetro aproximado de 400 m por lado, por lo que el número de unidades muestrales (transectos) fue relativamente bajo (N = 7 en total, tanto para invertebrados como para peces). Estas correspondieron a transectos de banda de 25 x 4 m, para peces (100 m2), y 25 x 2 m, para equinodermos (50 m2), los cuales se recorrieron de manera paralela a la costa, siguiendo la metodología estándar empleada en arrecifes del Pacífico mexicano (Fernández-Rivera, 2015; Meléndez-Rosas et al. 2023). Los transectos se colocaron siguiendo el relieve del fondo marino, por lo cual se establecieron a profundidades de entre 12 y 19 metros, de acuerdo con el punto de inicio, por medio de buceo autónomo. Se comenzó en la porción sur de la isla, recorriéndola, siempre siguiendo la línea de costa; cuatro transectos fueron colocados en dirección oeste y tres en la este. En cada transecto, la identificación de los especímenes observados se llevó a cabo con censos visuales, tomando como base las guías de Robertson & Allen (2015), Thomson et al. (2000) y Humann & DeLoach (2004), para peces, y de Gotshall (1987), Hickman (1998) y Kerstitch & Bertsch (2007), para erizos y estrellas de mar. Adicionalmente, se evaluó el fondo de estos sitios bajo las siguientes categorías: corales, arena, gravilla, grava, roca, bloque, tapete algal, alga filamentosa, alga verde, alga café, alga roja, alga incrustante coralina, alga articulada coralina, invertebrados y otros.
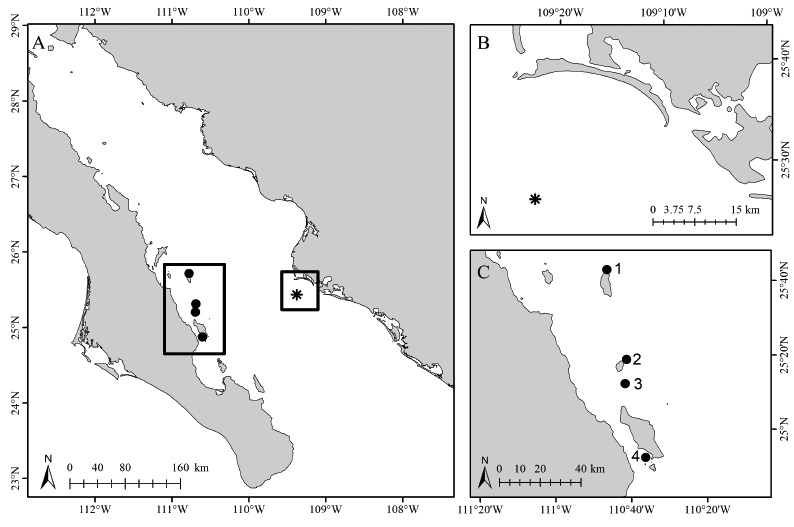
Fig. 1. Área de estudio: A) Sitios de estudio en el sur del golfo de California, México. B) Isla Farallón de San Ignacio (Sinaloa, México). C) Arrecifes rocosos insulares cercanos a la zona de estudio y analizados durante la investigación: 1) isla Catalana, 2) isla Santa Cruz, 3) isla San Diego, 4) El Cayo
Fig. 1. Study area. A) Study sites in the Southern Gulf of California, México. B) Main study site, Farallón de San Ignacio Island (Sinaloa, México). C) Insular rocky reefs close to the main study site that were also analyzed. 1) Catalana Island, 2) Santa Cruz Island, 3) San Diego Island, 4) El Cayo
Con los datos de los censos, se obtuvieron cuatro índices de estructura comunitaria: abundancia (N), riqueza (S), diversidad de Shannon-Wiener (H’) y equidad de Pielou (J’). Estos fueron analizados de manera descriptiva y comparados estadísticamente con los generados para cuatro lugares con fondos rocosos y presencia de corales, situados entre 130 y 140 km al oeste del Farallón de San Ignacio y cerca de la costa occidental del golfo de California: la islas Catalana, Santa Cruz, San Diego y El Cayo (Fig. 1C). Dichas localidades también fueron visitadas en noviembre de 2016 y en cada una se llevaron a cabo censos de peces e invertebrados, con la misma metodología, siempre de forma paralela a la costa.
Dado que no se cuenta con el mismo tamaño de muestra (transectos) en los analizados y que los datos no siempre cumplieron los requisitos de normalidad y homogeneidad de varianzas (Zar, 2010), se efectuaron análisis de una vía permutacionales, basados en matrices de distancias euclidianas (Anderson et al. 2008). Los análisis de varianza no paramétricos se realizaron con 10 000 permutaciones, siguiendo los criterios de Anderson et al. (2008) y con la definición del sitio como el factor fijo. Todas las pruebas estadísticas se realizaron con un valor alfa de 0.05.
Finalmente, para denotar el nivel de semejanza entre las faunas de peces e invertebrados de las cinco islas estudiadas, se efectuaron análisis de similitud (ANOSIM) (Clarke & Green, 1988) en el programa Primer ver. 6.0. (Clarke & Gorley, 2006). Este procedimiento se hizo separadamente por taxón y se basó en las abundancias de las especies presentes, con las cuales se construyeron matrices de similitud, empleando el coeficiente de Bray-Curtis para finiquitar los ANOSIM. Estos exámenes se lograron con 10 000 simulaciones.
Observaciones descriptivas. El listado de especies de peces, erizos y asteroideos observados en el Farallón de San Ignacio, junto con sus valores de densidad poblacional (individuos por metro cuadrado), se presenta en el cuadro 1. Se encontraron 34 especies de peces, dos de asteroideos y cuatro de equinoideos. Las especies dominantes de peces, en orden decreciente de abundancia fueron Azurina atrilobata Gill, 1862 (633 individuos), Prionurus laticlavius Gill, 1862 (79 individuos) y Microspathodon dorsalis Gill, 1862 (57 individuos). En equinoideos y asteroideos, los taxa más comunes fueron D. mexicanum (128 individuos) y P. unifascialis (55 individuos), respectivamente. Es importante mencionar que, en el caso de los asteroideos, en el Farallón de San Ignacio solo se encontraron en los transectos a P. unifascialis y P. pyramidata (Cuadro 1), aunque en los buceos se observó también a Acanthaster planci (Linnaeus, 1758), Mithrodia bradleyi (Verrill, 1870) y Pentaceraster cumingi (Gray, 1840).
Cuadro S1. Análisis a posteriori que evalúan las diferencias por pares entre las comunidades ícticas analizadas. El valor de significancia de cada prueba se muestra entre paréntesis; las pruebas significativas se exponen en negritas. S = riqueza, N = abundancia, EC = El Cayo, SD = San Diego, SC = Santa Cruz, EF = El Farallón, IC = isla Catalana
Table S1. A posteriori analyses evaluating pairwise differences among the analyzed fish communities. The significance value of each test is shown in parentheses; significant tests are shown in bold. S = richness, N = abundance, EC = El Cayo, SD = San Diego, SC = Santa Cruz, EF = El Farallón, IC = Catalana Island
|
Prueba t |
S |
N |
|
EC-SD |
2.067 (0.055) |
2.637 (0.006) |
|
EC-SC |
0.112 (0.912) |
0.307 (0.770) |
|
EC-EF |
1.046 (0.326) |
0.950 (0.383) |
|
EC-IC |
0.732 (0.479) |
0.264 (0.811) |
|
SD-SC |
1.846 (0.083) |
1.762 (0.093) |
|
SD-EF |
3.748 (0.001) |
3.155 (0.0003) |
|
SD-IC |
3.143 (0.006) |
3.816 (0.0009) |
|
SC-EF |
0.726 (0.489) |
0.995 (0.353) |
|
SC-IC |
0.486 (0.655) |
0.563 (0.580) |
|
EF-IC |
0.235 (0.819) |
0.754 (0.531) |
Análisis comunitario de las faunas del Farallón de San Ignacio y comparación con islas cercanas. Los valores promedio de riqueza (S), abundancia (N), diversidad (H’) y equidad (J’) para peces, asteroideos y equinoideos del Farallón, igual que de arrecifes situados en la misma latitud, pero en la costa oeste del golfo, se muestran en las figuras de la 2 a la 4; las pruebas a posteriori de los análisis con significancia estadística se pueden observar en el material suplementario 1 (cuadros S1-S3). Para peces, los valores promedio y el error típico de estos índices por unidad de censo en el Farallón fueron: riqueza: 13.71 ± 1.69; abundancia: 157.14 ± 51.84 ind/100m2; diversidad: 1.66 ± 0.20; y uniformidad: 0.66 ± 0.09 (Fig. 2). Los análisis estadísticos entre islas demostraron que los índices ecológicos de la comunidad íctica presentaron diferencias significativas en la riqueza (Pseudo-F = 3.123, P = 0.024. N = 97) y abundancia (Pseudo-F = 2.870, P = 0.030. N = 97), debido a que los valores fueron más bajos en isla Catalana que en isla San Diego (Fig. 2A, 2B), mas no difieren en la diversidad H’ (Pseudo-F = 0.211, P = 0.928. N = 97) ni en la equidad J’ (Pseudo-F = 0.426, P = 0.791. N = 97). En consecuencia, al no diferenciarse estadísticamente, puede decirse que el Farallón tuvo niveles de complejidad ecológica similares, en relación con los otros arrecifes insulares.
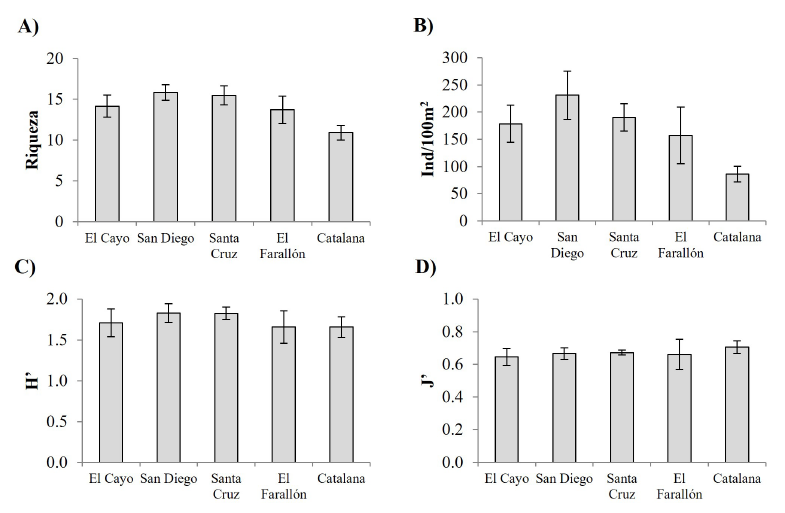
Fig. 2. Valores promedio (± error estándar) de índices ecológicos de la comunidad de peces observados en el Farallón de San Ignacio y en arrecifes rocosos situados en una latitud semejante. A) Riqueza, B) abundancia, C) diversidad, D) equidad
Fig. 2. Ecological indexes’ mean values (± standard error) from the fish community observed at Farallón de San Ignacio and other rocky reefs in similar latitudes. A) Species richness, B) abundance, C) diversity, D) evenness
Para asteroideos, los valores promedio de los índices por transecto en el Farallón fueron riqueza: 1.71 ± 0.33; abundancia: 9.43 ± 1.96 ind/100m2; diversidad: 0.14 ± 0.06; y equidad: 0.45 ± 0.16. En este taxón, se observaron diferencias significativas en la riqueza (Pseudo-F = 11.297, P < 0.0001), pues El Cayo presentó las cifras más altas (Fig. 3A). La abundancia (Pseudo-F = 19.151, P < 0.0001. N = 97) y la diversidad (Pseudo-F = 4.394, P = 0.003) fueron bajas en San Diego y Santa Cruz, diferenciándose por lo que se distinguen de los sitios restantes (Fig. 3B, 3C). Finalmente, la equidad fue similar entre todas las islas (Pseudo-F = 2.033, P = 0.095) (Fig. 3D). Por lo tanto, el Farallón no presentó diferencias significativas en la complejidad estructural de los asteroideos, al contrastar con los demás arrecifes insulares.
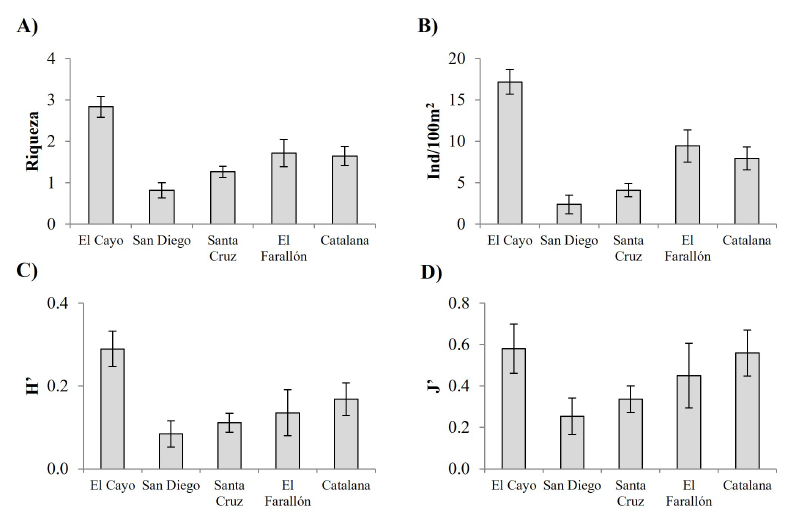
Fig. 3. Valores promedio (± error estándar) de índices ecológicos de las comunidades de asteroideos observados en el Farallón de San Ignacio (Sinaloa, México) y en arrecifes rocosos situados en una latitud semejante. A) Riqueza, B) abundancia, C) diversidad, D) equidad
Fig. 3. Ecological indexes’ mean values (± standard error) from the asteroidea community observed at Farallón de San Ignacio and other rocky reefs in similar latitudes. A) Species richness, B) abundance, C) diversity, D) evenness
Respecto a los equinoideos, los valores promedio de los índices en el Farallón fueron riqueza: 2.71 ± 0.34; abundancia: 34.18 ± 2.93 ind/100m2; diversidad: 0.33 ± 0.05; y uniformidad: 0.76 ± 0.15. A diferencia de lo observado en peces y asteroideos, en el caso de los equinoideos el Farallón destacó por presentar los índices ecológicos con los valores más altos (Fig. 4), por lo que sobresale, claramente, del resto de los sitios y presenta abundancias hasta diez veces mayores que las vistas en San Diego y Santa Cruz. En consecuencia, se notaron diferencias significativas en todos los índices: riqueza (Pseudo-F = 6.363, P = 0.001), abundancia (Pseudo-F = 27.131, P < 0.0001, N = 97), diversidad (Pseudo-F = 5.924, P = 0.0004) y equidad (Pseudo-F = 3.209, P = 0.014), lo cual indica que el Farallón presentó la comunidad más rica, abundante y diversa de erizos de mar entre los lugares revisados (Fig. 4). Esto es importante, considerando que, en comparación con otros sitios, en el Farallón de San Ignacio las unidades muestrales fueron menores; destaca la trascendencia de este punto geográfico, ya que, en el resto, con mayores unidades muestrales, los índices ecológicos obtuvieron valores más bajos.
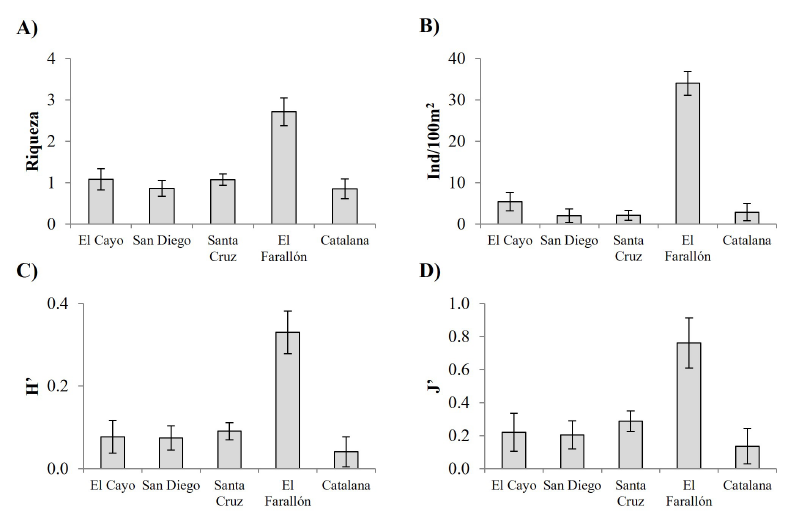
Fig. 4. Valores promedio (± error estándar) de índices ecológicos de las comunidades de equinoideos observados en el Farallón de San Ignacio (Sinaloa, México) y en arrecifes rocosos situados en una latitud semejante. A) Riqueza, B) abundancia, C) diversidad, D) equidad
Fig. 4. Ecological indexes mean values (± standard error) from the echinoidea community observed at the Farallón de San Ignacio and other rocky reefs in similar latitudes. A) Species richness, B) abundance, C) diversity, D) evenness
El análisis global de similitud indica que, por su composición de especies (fauna total) y sus abundancias relativas, todos los sitios son semejantes entre sí (R = 0.393). Sin embargo, al momento de hacer las comparaciones entre pares de lugares, el Farallón se diferenció notablemente de los demás (valores de R mayores a 0.6; Cuadro S4), mientras que el resto de las zonas presentaron las comunidades más similares entre sí. Esto mismo ocurre al comparar los peces (R = 0.354) y erizos (R = 0.219), mientras que en el grupo de los asteroideos existe cierto nivel de diferenciación entre las áreas analizadas (R = 0.588); el Farallón y el Cayo mostraron el mayor nivel de diferenciación (ver material suplementario).
La mayoría de las investigaciones realizadas en el Farallón de San Ignacio se enfocan en aves y mamíferos marinos presentes en esta isla. Este es el primer estudio que analiza en conjunto la estructura comunitaria de peces y equinodermos asociados a zonas rocosas de la costa del estado de Sinaloa, México. En ese sentido, no es posible hacer comparaciones con resultados obtenidos en hábitats semejantes en la misma región, particularmente, por el gran número de investigaciones históricas realizadas, por ejemplo, en la costa occidental del golfo. El único trabajo análogo es el de Caso-Muñoz et al. (1996), efectuado en la bahía de Mazatlán, en el cual se calcularon índices ecológicos de riqueza y diversidad para equinodermos. Sin embargo, el trabajo se hizo a lo largo de casi una década de visitas a la zona de estudio, se examinó la fauna en su conjunto (las cuatro clases acumuladas) y se incluyeron especies de fondos blandos y duros; por estas razones, es complejo llevar a cabo comparaciones consistentes, en especial, cuando se aborda más de un grupo en la presente indagación. Debido a las discrepancias anteriores que limitan los análisis posibles y dada la relevancia de llenar este vacío de información, se tomaron en cuenta solo los datos recolectados durante la misma campaña de muestreo en arrecifes rocosos que cuentan con las mismas condiciones topográficas, ambientales y que, además, se encontraran en latitudes similares. Esto con el fin de que la información obtenida fuera comparable y se redujera el sesgo en los análisis estadísticos, provocado por cambios en latitud y diferencias de microhábitats.
A pesar de las limitaciones mencionadas, pueden discutirse diversos patrones vistos en los resultados entre los sitios seleccionados: los análisis estadísticos y de similitud denotaron diferencias significativas que logran separar al Farallón de San Ignacio del resto de los arrecifes insulares evaluados. Tal es el marco de referencia que se toma para esta sección. Cabe mencionar que dichos arrecifes rocosos fueron elegidos por encontrarse en la misma latitud que el Farallón y por presentar profundidades, condiciones ambientales y topografías semejantes, por lo cual se esperaría que mostraran estructuras comunitarias y una composición faunística semejantes.
La primera observación relevante es que las tres especies de peces dominantes en el Farallón (A. atrilobata, P. laticlavius y M. dorsalis) tienen diferencias notables de tamaño y hábitos alimenticios entre ellas. Según datos referidos por Robertson & Allen (2015) y Froese & Pauly (2018), la primera alcanza una longitud total de hasta 13 cm y es consumidora de plancton, mientras que el cirujano P. laticlavius es mucho mayor (hasta 60 cm de longitud total) y funge como herbívoro. Por último, la damisela M. dorsalis presenta una talla intermedia en relación con las otras dos especies (hasta 31 cm de longitud total), es una especie demersal y sus hábitos son omnívoros. También es importante mencionar que A. atrilobata y P. laticlavius no son territoriales, mientras que M. dorsalis sí, aunque únicamente en temporada reproductiva (Thomson et al. 2000). Todas estas diferencias en conjunto indican que las especies, posiblemente, no sean cercanas en lo referente a su nicho ecológico, lo que contribuye a que puedan coexistir con altos números, al no presentar competencia directa o intensa por los recursos.
Por otra parte, con excepción de la damisela punto blanco A. atrilobata, las otras especies de peces más abundantes del Farallón no corresponden a las consideradas como dominantes en estudios previos sobre la ictiofauna arrecifal del sur del golfo de California (Pérez-España et al. 1996; Aburto-Oropeza & Balart, 2001; Arreola-Robles & Elorduy-Garay, 2002; Rodríguez-Romero et al. 2005; Álvarez-Filip et al. 2006; Barjau-González et al. 2012). Todos los trabajos citados señalan a A. atrilobata, junto a Thalassoma lucasanum (Gill, 1862) y Stegastes rectifraenum (Gill, 1862), como los peces más abundantes en los arrecifes entre cabo Pulmo (23°N) e isla San José (25°N). Es difícil explicar la causa de esta diferencia entre islas de los dos márgenes del golfo. No obstante, como hipótesis, lo observado puede ligarse al pequeño tamaño del Farallón de San Ignacio, que no ofrece opciones para colocar numerosos territorios de S. rectifraenum, los cuales son permanentes (Thomson et al. 2000), o bien el hábitat quizá no es el adecuado, ya que el fondo es poco heterogéneo y la densidad de corales es muy baja en comparación con el resto de los sitios analizados (menos del 3% de cobertura de coral allí). Esto no favorece la presencia del lábrido T. lucasanum ni brinda protección para S. rectifraenum, asociado a fondos con formaciones rocosas de tipo escombro, que no predominan en el Farallón (Aburto-Oropeza & Balart, 2001; Álvarez-Filip et al. 2006). Adicionalmente, la dominancia de P. laticlavius y M. dorsalis apunta a la disponibilidad del recurso algal como parte de la dieta. En este sentido, datos sobre la cobertura del fondo de la zona de estudio generados en paralelo con los censos de peces e invertebrados muestran una cobertura algal del 42.3%, dominada por las algas coralinas incrustantes (20.4%) y los tapetes de cianofitas (15.9%). La ocurrencia de estos dos tipos algales ilustra intensidades altas de herbivorismo (Birkeland, 2015), en este caso efectuado por los peces citados y los erizos, cuya abundancia y riqueza fue muy elevada.
El número relativamente disminuido de especies de peces registrado en Farallón (34 en total) ocasionó que el sitio apareciera como relativamente atípico en relación con otras islas de la misma latitud. La baja riqueza se puede interpretar como resultado de que el esfuerzo de muestreo no fue muy alto (bajo número de réplicas), sin embargo, es necesario recordar que este estudio es una primera aproximación y que se cubrió el área mínima necesaria de monitoreo recomendada para arrecifes rocosos del golfo de California (Fernández-Rivera Melo, 2015). Esta es la metodología estándar aplicada en arrecifes rocosos y coralinos del Pacífico mexicano (Meléndez-Rosas et al. 2023). Otra interpretación puede ser el pequeño tamaño del área en sí, en contraste con las otras analizadas; pero, este último factor y el hecho de que la riqueza ictiofaunística en el golfo de California está ligada a la superficie de la plataforma insular (Thomson & Gilligan, 2002) pueden hacer que el resultado obtenido tenga una explicación ecológica, ligada a aspectos de la relación área-especies. Este patrón se repite en múltiples tipos de organismos y en franjas tanto marinas como terrestres (Parravicini et al. 2013). Asimismo, de manera sucinta, demuestra que a mayor tamaño de una zona insular marina o parche de algún tipo particular de hábitat (en el caso terrestre), más grande será el número de especies que lo ocupen. La mejor explicación para la existencia de la relación área-especies en organismos acuáticos es que, en una zona geográfica dada, la superficie determina el número potencial de hábitats presentes (Thomson & Gilligan, 2002). Si este es el caso, en el Farallón de San Ignacio la cantidad de hábitats submareales disponibles debe ser reducida, lo que se debe traducir en una baja diversidad gama en los peces, comparada con las otras islas revisadas.
A pesar de lo anterior, la riqueza, abundancia, diversidad y equidad de peces por unidad de muestreo (transectos) en el Farallón no presentaron diferencias significativas con los otros arrecifes insulares revisados, que se encuentran en latitudes similares. Eso indica que, desde la perspectiva estructural, los ensamblajes de esta localidad están a la par de los otros y apunta hacia el hecho de que, a pesar de su localización geográfico y relativo aislamiento, la zona de estudio mantiene una complejidad ecológica alta o, al menos, promedio. Sería importante llevar a cabo análisis de diversidad funcional, empleando metodologías y clasificaciones ya disponibles para el Pacífico mexicano (Olivier et al. 2018; Ramírez-Ortiz et al. 2018; Dubuc et al. 2023), con el propósito de confirmar si esta condición también se mantiene.
En cuanto a los asteroideos, tanto P. unifascialis como P. pyramidata están reconocidas como las más abundantes de este grupo taxonómico a lo largo del golfo de California, tanto en zonas insulares como costeras (Caso-Muñóz et al. 1996; Reyes-Bonilla et al. 2005; González-Medina et al. 2006; Holguín-Quiñones et al. 2008; Luna-Salguero & Reyes-Bonilla, 2010; Ulate et al. 2016) y su ubicua presencia en arrecifes de esa región se ha explicado por la gran abundancia de tapetes de algas que son parte de su alimento principal (Cintra-Buenrostro et al. 2005). En este sentido, la zona de estudio se comporta típicamente en lo que se refiere a los arrecifes rocosos del golfo.
No obstante, es importante mencionar que el Farallón presentó un muy bajo número de taxones, al compararlo con otras regiones, y esto hizo que el análisis de ordenamiento lo diferenciara claramente del resto. En áreas como La Paz (24°N), Loreto (25°N) o Bahía de Los Ángeles (28°N), en estudios que involucran censos en el campo (Reyes-Bonilla et al. 2005; Luna-Salguero & Reyes-Bonilla, 2010; Herrero-Pérezrul et al. 2015; Ulate et al. 2016), se reportan entre seis y nueve especies, además de se citan, como relativamente frecuentes y abundantes, otras estrellas como A. planci, M. bradleyi y P. cumingi, las cuales también se contabilizaron en las islas de la costa occidental del golfo en el 2016 y en el Farallón, pero en números muy bajos, de manera que no aparecieron en las unidades de muestreo. Esto último puede deberse a situaciones de preferencia de hábitat; por ejemplo, P. cumingi tiene predilección por fondos blandos (Reyes-Bonilla et al. 2016) y la falta de corales ayudar a explicar la poca densidad de A. planci, ya que los cnidarios representan una fuente importante de alimento para la especie. Sin embargo, la homogeneidad del fondo en el Farallón y el pequeño tamaño de la isla también pueden tener un efecto depresor de la riqueza de los asteroideos.
Finalmente, para los erizos de mar las diferencias no se presentan en la composición, ya que las cuatro especies observadas en el Farallón son comunes en los arrecifes rocosos del golfo de California (Holguín-Quiñones et al. 2000; González-Medina et al. 2006; Herrero-Pérezrul et al. 2007; Holguín-Quiñones et al. 2008). Al mismo tiempo, se vio en la zona a Echinometra vanbrunti A. Agassiz, 1863, aunque en aguas someras donde no se llevaron a cabo muestreos. No obstante, al comparar con los otros sitios de estudio, se observó que el Farallón posee densidades muy altas de equinoideos, en especial de D. mexicanum. Esto resulta interesante, ya que ese erizo no es en extremo abundante en el golfo de California (Alvarado et al. 2016) y sus densidades en el Farallón se asemejan más a lo reportado en arrecifes del Pacífico central y sur de México, donde es dominante (Honey-Escandón et al. 2008; Ríos-Jara et al. 2013; Solís-Marín et al. 2014; Rojas-Montiel & Benítez-Villalobos, 2015; Hermosillo-Nuñez et al. 2016). La notable abundancia de D. mexicanum también explica la diferencia en composición entre el Farallón y las demás islas, en el análisis de similitud, y es congruente con las observaciones sobre la gran cantidad de algas coralinas incrustantes en el fondo, ya que este organismo es bioerosionador (Alvarado et al. 2015).
Este estudio presentó la primera descripción de los ensamblajes de peces, asteroideos y equinoideos del Farallón de San Ignacio, el único arrecife rocoso insular en el sureste del golfo de California localizado dentro de la llamada “brecha faunística de Sinaloa”. Los resultados evidenciaron que la complejidad de la estructura comunitaria no difiere de otras islas del suroeste del golfo, ya que no hubo diferencias significativas en riqueza, abundancia, diversidad o equidad. No obstante, la composición de especies fue sensiblemente distinta a la observada en islas de Baja California Sur, debido, en especial, a la gran incidencia de especies herbívoras en el Farallón, la cual se explica por su alta cobertura algal.
El Consejo Nacional de Ciencia y Tecnología de México (proyecto Fronteras en la Ciencia clave 292, a HRB) aportó los recursos para realizar el trabajo de campo. Se agradece a Amy Hudson Weaver y Ollin González Cuéllar (Sociedad de Historia Natural Niparajá), por ceder la información sobre la fauna marina presente en las islas del occidente del golfo de California, la cual fue generada con recursos de la David and Lucile Packard Foundation, The Walton Family Foundation, The Nature Conservancy, el Fondo Mexicano para la Conservación de la Naturaleza, más La Alianza WWF y la Fundación Carlos Slim. Rebeca Granja (Universidad Autónoma Metropolitana, Iztapalapa) y Luis Hernández (UABCS) amablemente revisaron versiones preliminares de la nota. Durante la preparación del documento, MOG y VMC fueron receptores de becas del Consejo Nacional de Ciencia y Tecnología (registro 413585 y 332939, respectivamente). De igual forma, los autores agradecen los comentarios de tres evaluadores anónimos que mejoraron el manuscrito.
Aburto-Oropeza, O. & Balart, E. F. (2001). Community structure of reef fish in several habitats of a rocky reef in the Gulf of California. Mar. Ecol., 22(4), 283-305. https://doi.org/10.1046/j.1439-0485.2001.01747.x
Alvarado, J. J., Reyes-Bonilla, H. & Benítez-Villalobos, F. (2015). Diadema mexicanum, erizo de mar clave en los arrecifes coralinos del Pacífico Tropical Oriental: lo que sabemos y perspectivas futuras (Diadematoida: Diadematidae). Rev. Biol. Trop., 63(2), 135-157.
Alvarado, J. J., Cortés, J., Guzmán, H. & Reyes-Bonilla, H. (2016). Density, size, and biomass of Diadema mexicanum (Echinoidea) in Eastern Tropical Pacific coral reefs. Aquat. Biol., 24 (3), 151-161. https://doi.org/10.3354/ab00645
Álvarez-Filip, L., Reyes-Bonilla, H. & Calderón-Aguilera, L. E. (2006). Community structure of fishes in Cabo Pulmo reef, Gulf of California. Mar. Ecol., 27(3), 253-262. https://doi.org/10.1111/j.1439-0485.2006.00108.x
Anderson, M. J., Gorley R. N. & Clarke, K. R. (2008). PERMANOVA+ for PRIMER: guide to software and statistical methods. United Kingdom. PRIMER-E Press.
Arreola-Robles, J. L. & Elorduy-Garay, J. F. (2002). Reef fish diversity in the region of La Paz, Baja California Sur, Mexico. Bull. Mar. Sci., 70(1), 1-18.
Barjau-González, E., Rodríguez-Romero, J., Galván-Magaña, F. & López-Martínez, J. (2012). Changes in the taxonomic diversity of the reef fish community of San José Island, Gulf of California, Mexico. Biodivers. Conserv., 21(14), 3543-3554. https://doi.org/10.1007/s10531-012-0378-z
Birkeland, C. (2015). Geographic differences in ecological processes on coral reefs. In C. Birkeland (Ed.), Coral Reefs in the Anthropocene. (pp. 179-194). Springer, Holland. https://doi.org/10.1007/978-94-017-7249-5_9
Brusca, R. C. (1980). Common intertidal invertebrates of the Gulf of California. EE.UU. The University of Arizona Press.
Caso-Muñóz, M. E., Laguarda-Figueras, A., Solís-Marín, F. A., Ortega-Salas, A. & Durán-González, A. de la Luz. (1996). Contribución al conocimiento de la ecología de las comunidades de equinodermos de la Bahía de Mazatlán, Sinaloa, México. An. Inst. Cienc. Mar Limnol. Univ. Nac. Autón. Méx., 22(1), 101-119.
Cintra-Buenrostro, C. E., Reyes-Bonilla, H. & Herrero-Pérezrul, M. D. (2005). Oceanographic conditions and diversity of sea stars (Echinodermata: Asteroidea) in the Gulf of California, Mexico. Rev. Biol. Trop., 53(3), 245-261.
Clarke, K. R. & Green, R. H. (1988). Statistical design and analysis for a ’biological effects’ study. Mar. Ecol. Progr. Ser., 46, 213-226.
Clarke, K. R. & Gorley, R. N. (2006). PRIMER v6: user manual/tutorial (Plymouth routines in multivariate ecological research). United Kingdom. Primer-E Ltd.
Cupul-Magaña, A. L. (2003). Comunidades coralinas de Sinaloa. En L. J. L. Cifuentes & J. Gaxiola (Eds.), Atlas de los ecosistemas de Sinaloa. (pp. 91-97). El Colegio de Sinaloa, Culiacán.
Dubuc, A., Quimbayo J. P., Alvarado, J. J., Araya-Arce, T., Arriaga, A., Ayala-Bocos, A., ... & Bejarano, S. (2023). Patterns of reef fish taxonomic and functional diversity in the Eastern Tropical Pacific. Ecography, 10: e06536. https://doi.org/10.1111/ecog.06536
Fernández-Rivera Melo, F. J. (2015). Análisis de los métodos de monitoreo empleados para evaluar el estado de las Áreas Naturales Protegidas marinas en el Golfo de California. (Tesis de maestría no publicada). Universidad Autónoma de Baja California Sur.
Froese, R. & Pauly, D. (2018). FishBase. World Wide Web electronic publication. http://www.fishbase.org
González-Bernal, M. A., Mellink, E. & Fong-Mendoza, J. R. (2002). Nesting birds of Farallón de San Ignacio, Sinaloa, México. West. Birds, 33(4), 254-257.
González-Medina, F. J., Holguín-Quiñones, O. E. & De la Cruz-Agüero, G. (2006). Variación espaciotemporal de algunos macroinvertebrados (Gastropoda, Bivalvia y Echinodermata) de fondos someros del Archipiélago Espíritu Santo, Baja California Sur, México. Cienc. Mar., 32(1A), 33-44. https://doi.org/10.7773/cm.v32i1.67
Gotshall, D. W. (1987). Marine Animals of the Baja California: a guide to the common fishes and invertebrates. EE. UU. Sea Challengers Inc.
Granja-Fernández, R., Maya-Alvarado, B., Rodríguez-Zaragoza, F. A. & López-Pérez, A. (2023). Ophiuroidea (Echinodermata) diversity partitioning across the Eastern tropical Pacific. Reg. Stud. Mar. Sci., 60, 102835. https://doi.org/10.1016/j.rsma.2023.102835
Guevara-Medina, M. A., Castillo-Guerrero, J. A. & González-Bernal, M. A. (2008). Presencia y abundancia de aves de la isla Farallón de San Ignacio, Sinaloa. Huitzil, 9(2), 20-28.
Hastings, P. A. (2000). Biogeography of the Tropical Eastern Pacific: distribution and phylogeny of chaenopsid fishes. Zool. J. Linn. Soc., 128(3), 319-335. https://doi.org/10.1111/j.1096-3642.2000.tb00166.x
Hermosillo-Núñez, B. B., Rodríguez-Zaragoza, F. A., Ortiz, M., Calderón-Aguilera, L. E. & Cupul-Magaña, A. L. (2016). Influence of the coral reef assemblages on the spatial distribution of echinoderms in a gradient of human impacts along the tropical Mexican Pacific. Biodivers. Conserv., 25(11), 2137-2152. https://doi.org/10.1007/s10531-016-1182-y
Herrero-Pérezrul, M. D., Reyes-Bonilla, H., González-Azcárraga, A., Cintra-Buenrostro, C. E. & Rojas-Sierra, A. (2007). Equinodermos. En G. D. E. Danemann & E. D. Exequiel (Eds.), Bahía de los Ángeles: Recursos Naturales y Comunidad, línea Base 2007 (pp. 339-357). México. Secretaría de Medio Ambiente y Recursos Naturales / Instituto Nacional de Ecología / PRONATURA Noroeste, San Diego Natural History Museum.
Herrero-Pérezrul, M. D., Rojero-León, S. & Reyes Bonilla, H. (2009). Community structure of conspicuous echinoderms of three islands from the Gulf of California, Mexico. In L. G. Harris, S. A. Boettger, C. W. Walker & M. P. Lesser (Eds.), Echinoderms: Durham (pp. 615-620). Proccedings of the 12th International Echinoderm Conference
Herrero-Pérezrul, M. D., Ramírez-Ortiz, G., Rosales-Estrada, M. & Reyes-Bonilla, H. (2015). Densidad poblacional y distribución espacial de erizos de mar (Echinodermata: Echinoidea) en la Isla Socorro, Archipiélago de Revillagigedo, México. Rev. Biol. Trop., 63(2), 221-232.
Hickman, C. P. (1998). A field guide to sea stars & other echinoderms of Galapagos (Galapagos Marine Life Series). EE. UU. Sugar Spring Press.
Holguin-Quiñones, O., Wright-López, H. & Solís-Marín, F. (2000). Asteroidea, Echinoidea y Holothuroidea en fondos someros de la Bahía de Loreto, Baja California Sur, México. Rev. de Biol. Trop., 48(4), 749-757.
Holguin-Quiñones, O. E., González-Medina, F. J., Solís-Marín, F. & Félix-Pico, E. F. (2008). Variación espacio-temporal de Scleractinia, Gorgonacea, Gastropoda, Bivalvia, Cephalopoda, Asteroidea, Echinoidea y Holothuroidea, de fondos someros de la isla San José, Golfo de California. Rev. Biol. Trop., 56(3), 1189-1199. https://doi.org/10.15517/rbt.v56i3.5703
Honey-Escandón, M., Solís-Marín, F. A. & Laguarda-Figueras, A. (2008). Equinodermos (Echinodermata) del Pacífico Mexicano. Rev. Biol. Trop., 56(3), 57-73.
Humann, P. & Deloach, N. (2004). Reef Fish Identification: Baja to Panama. EE. UU.: New World Publications, Jacksonville.
Kerstitch, A. & Bertsch, H. (2007). Sea of Cortez Marine Invertebrates. 2nd ed. EE.UU.: Sea Challengers, Monterrey.
Luna-Salguero, M. B. & Reyes Bonilla, H. (2010). Estructura comunitaria y trófica de las estrellas de mar (Echinodermata: Asteroidea) en arrecifes rocosos de Loreto, Golfo de California, México. Hidrobiológica, 20(2), 127-134.
Maya-Alvarado, B., Granja-Fernández, R., López-Pérez, A. & Rodríguez-Zaragoza, F. A. (2023). Diversity patterns of echinoids (Echinodermata: Echinoidea) in shallow waters of the Mexican Pacific. Reg. Stud. Mar. Sci., 68, 103246. https://doi.org/10.1016/j.rsma.2023.103246
Medina-Rosas, P. (2006). Los corales hermatípicos (Anthozoa: Scleractinia) de Mazatlán, Sinaloa. Cien. Mar., 10(28), 13-17.
Meléndez-Rosas, R., García-Vázquez, G., Franco-Ortíz, M., Reyes-Bonilla, H., Cupul-Magaña, A., López-Pérez, A., … & Saldierna-Calápiz, D. (2023). Fortalecimiento del manejo y conservación de corales en Áreas Naturales Protegidas del Pacífico mexicano mediante una red colaborativa. Hidrobiológica 33(2), 265-271. https://doi.org/10.24275/FXIH7586
Olivier, D., Loiseau, N., Petatán-Ramírez, D., Trujillo-Millán, O., Suárez-Castillo, A. N., Torre, J., Munguía-Vega, A. & Reyes-Bonilla, H. (2018). Functional-biogeography of the reef fishes of the islands of the Gulf of California: Integrating functional divergence into marine conservation. Glob. Ecol. Conserv., 16, e00506. https://doi.org/10.1016/j.gecco.2018.e00506
Parravicini, V., Kulbicki, M., Bellwood, D. R., Friedlander, A. M., Arias-González, J. E., Chabanet, P., ... & Mouillot, D. (2013). Global patterns and predictors of tropical reef fish species richness. Ecography, 36(12), 1254-62. https://doi.org/10.1111/j.1600-0587.2013.00291.x
Pérez-España, H., Abitia-Cárdenas, L. A. & Galván-Magaña, F. (1996). Variaciones temporales y espaciales en la estructura de la comunidad de peces de arrecifes rocosos del suroeste del Golfo de California, México. Cienc. Mar., 22(3), 273-294. https://doi.org/10.7773/cm.v22i3.864
Ramírez-Ortiz, G., Calderón-Aguilera, L. E., Reyes-Bonilla, H., Ayala-Bocos, A., Hernández, L., Fernández Rivera-Melo, F., … & Dominici-Arosamena, A. (2018). Functional diversity off fish and invertebrates in coral and rocky reefs of the Eastern Tropical Pacific. Mar. Ecol., 38 (4), e12447. https://doi.org/10.1111/maec.12447
Reyes Bonilla, H., González-Azcárraga, A. & Rojas-Sierra, A. (2005). Estructura de las asociaciones de las estrellas de mar (Asteroidea) en arrecifes rocosos del Golfo de California, México. Rev. Biol. Trop., 53(3), 233-244.
Reyes-Bonilla, H. & López-Pérez, R. A. (2009). Corals and coral reef communities in the Gulf of California. In A. Johnson & J. Ledezma-Vázquez (Eds.), Atlas of coastal ecosystems in the western Gulf of California (pp. 45-57). The University of Arizona Press, Tucson. https://doi.org/10.2112/10A-00010.1
Reyes-Bonilla, H., Vázquez-Arce, D., González-Cuéllar, O. T., Herrero-Pérezrul, M. D. & Weaver, A. H. (2016). Mass aggregation of the cushion starfish Pentaceraster cumingi in the southern Gulf of California. Mar. Biodivers., 48(2), 949-950. https://doi.org/10.1007/s12526-016-0529-7
Ríos-Jara, E., Galván-Villa, C. M., Rodríguez-Zaragoza, F. A., López-Uriarte, E., Bastida-Izaguirre, D. & Solís-Marín, F. A. (2013). Los equinodermos (Echinodermata) de bahía Chamela, Jalisco, México. Rev. Mex. Biodivers., 84(1), 263-279. https://doi.org/10.7550/rmb.30461
Robertson, D. R. & Allen, G. R. (2015). Shore fishes of the tropical eastern Pacific: online information system. Version 2.0. http://biogeodb.stri.si.edu/sftep/
Robertson, D. R. & Cramer, K. L. (2009). Shore fishes and biogeographic subdivisions of the Tropical Eastern Pacific. Mar. Ecol. Prog. Ser., 380, 1-17. https://doi.org/10.3354/meps07925
Rodríguez-Romero, J., Muhlia-Melo, A. F., Galván-Magaña, F., Gutiérrez-Sánchez, F. J. & García-López, V. (2005). Fish assemblages around Espíritu Santo island and Espíritu Santo seamount in the lower Gulf of California, México. Bull. Mar. Sci., 77(1), 33-50.
Rojas-Montiel, B. & Benítez-Villalobos, F. (2015). Ecología poblacional de Diadema mexicanum (Echinodermata: Echinoidea) en dos comunidades arrecifales de bahías de Huatulco, Oaxaca, México. Rev. Mex. Biodivers., 86(1) 123-130. https://doi.org/10.7550/rmb.42918
Samaniego-Herrera, A., Aguirre-Muñoz, A., Félix-Lizárraga, M., Valdez-Villavicencio, J., González-Gómez, R., Rodríguez-Malagón, M., …. & Tershy, B. R. (2009). Eradication of black rats from Farallón de San Ignacio and San Pedro Mártir islands. In C. C. Damiani & D. K. Garcelon (Eds.), Proceedings of the 7th California Islands Symposium (pp. 337-347). EE. UU.: Institute for Wildlife Studies, Arcata.
Solís-Marín, F. A., Laguarda-Figueras, A. & Honey-Escandón, M. (2014). Biodiversidad de equinodermos (Echinodermata) en México. Rev. Mex. Biodivers., 85(1), 441-449. https://doi.org/10.7550/rmb.31805
Thomson, D. A., Findley, L. T. & Kerstitch, A. N. (2000). Reef fishes of the Sea of Cortez: the rocky-shore fishes of the Gulf of California. EE. UU.: University of Texas Press. https://doi.org/10.1086/411794
Thomson, D. A. & Gilligan, M. R. (2002). Rocky-shore fishes. In T. J. M. L. Cody & E. Ezcurra (Eds.), A new island biogeography of the Sea of Cortés (pp. 154-180). United Kingdom: Oxford University Press. https://doi.org/10.1086/421674
Ulate, K., Sánchez, C., Sánchez-Rodríguez, A., Alonso, D., Aburto-Oropeza, O. & Huato-Soberanis, L. (2016). Latitudinal regionalization of epibenthic macroinvertebrate communities on rocky reefs in the Gulf of California. Mar. Biol. Res., 12(4), 389-401. https://doi.org/10.1080/17451000.2016.1143105
Zar, J. H. (2010). Biostatistical Analysis. EE. UU. Prentice Hall. Dorling Kindersley.
Se muestran las pruebas a posteriori, para evaluar las diferencias entre pares de sitios en los índices ecológicos analizados, únicamente de aquellos análisis de varianza de una vía, que mostraron diferencias significativas (cuadros S1-S3). De igual forma, se exponen los valores de R en las comparaciones pareadas, que derivan de los análisis de similitud ANOSIM (cuadro S4)
Cuadro S1. Análisis a posteriori que evalúan las diferencias por pares entre las comunidades ícticas analizadas. El valor de significancia de cada prueba se muestra entre paréntesis; las pruebas significativas se exponen en negritas. S = riqueza, N = abundancia, EC = El Cayo, SD = San Diego, SC = Santa Cruz, EF = El Farallón, IC = isla Catalana
Table S1. A posteriori analyses evaluating pairwise differences among the analyzed fish communities. The significance value of each test is shown in parentheses; significant tests are shown in bold. S = richness, N = abundance, EC = El Cayo, SD = San Diego, SC = Santa Cruz, EF = El Farallón, IC = Catalana Island
|
Prueba t |
S |
N |
|
EC-SD |
2.067 (0.055) |
2.637 (0.006) |
|
EC-SC |
0.112 (0.912) |
0.307 (0.770) |
|
EC-EF |
1.046 (0.326) |
0.950 (0.383) |
|
EC-IC |
0.732 (0.479) |
0.264 (0.811) |
|
SD-SC |
1.846 (0.083) |
1.762 (0.093) |
|
SD-EF |
3.748 (0.001) |
3.155 (0.0003) |
|
SD-IC |
3.143 (0.006) |
3.816 (0.0009) |
|
SC-EF |
0.726 (0.489) |
0.995 (0.353) |
|
SC-IC |
0.486 (0.655) |
0.563 (0.580) |
|
EF-IC |
0.235 (0.819) |
0.754 (0.531) |
Cuadro S2. Análisis a posteriori que evalúan las diferencias por pares entre las comunidades de asteroideos analizadas. El valor de significancia de cada prueba se muestra entre paréntesis; las pruebas significativas se exponen en negritas. S = riqueza, N = abundancia, H’ = diversidad, EC = El Cayo, SD = San Diego, SC = Santa Cruz, EF = El Farallón, IC = isla Catalana
Table S2. A posteriori analyses assessing pairwise differences among the asteroid communities analyzed. The significance value of each test is shown in parentheses; significant tests are shown in bold. S = richness, N = abundance, H’ = diversity, EC = El Cayo, SD = San Diego, SC = Santa Cruz, EF = El Farallón, IC = Catalana Island
|
Prueba t |
S |
N |
H’ |
|
EC-SD |
5.711 (<0.0001) |
7.648 (<0.0001) |
3.666 (0.002) |
|
EC-SC |
5.116 (<0.0001) |
7.069 (<0.0001) |
3.523 (0.001) |
|
EC-EF |
2.681 (0.033) |
2.31 (0.036) |
2.152 (0.047) |
|
EC-IC |
3.843 (0.001) |
3.343 (0.003) |
2.03 (0.056) |
|
SD-SC |
1.817 (0.092) |
1.517 (0.136) |
0.685 (0.51) |
|
SD-EF |
2.358 (0.037) |
4.517 (0.0005) |
0.846 (0.391) |
|
SD-IC |
2.966 (0.007) |
3.608 (0.006) |
1.723 (0.97) |
|
SC-EF |
1.274 (0.249) |
2.726 (0.01) |
0.408 (0.691) |
|
SC-IC |
1.485 (0.199) |
2.422 (0.017) |
1.264 (0.212) |
|
EF-IC |
0.312 (1) |
0.574 (0.61) |
0.568 (0.573) |
Cuadro S3. Análisis a posteriori que evalúan las diferencias por pares entre las comunidades de equinoideos analizadas. El valor de significancia de cada prueba se muestra entre paréntesis; las pruebas significativas se exponen en negritas. S = riqueza, N = abundancia, H’ = diversidad, J’ = equidad, EC = El Cayo, SD = San Diego, SC = Santa Cruz, EF = El Farallón, IC = isla Catalana
Table S3. A posteriori analyses evaluating pairwise differences among the analyzed echinoid communities. The significance value of each test is shown in parentheses; significant tests are shown in bold. S = richness, N = abundance, H’ = diversity, J’ = evenness, EC = El Cayo, SD = San Diego, SC = Santa Cruz, EF = El Farallón, IC = Catalana Island
|
Prueba t |
S |
N |
H’ |
J’ |
|
EC-SD |
0.551 (0.645) |
2.212 (0.031) |
0.005 (0.947) |
0.111 (0.894) |
|
EC-SC |
0.004 (1) |
3.352 (0.003) |
0.291 (0.788) |
0.476 (0.567) |
|
EC-EF |
4.396 (0.001) |
3.551 (0.0002) |
3.967 (0.003) |
3.221 (0.006) |
|
EC-IC |
0.736 (0.619) |
1.647 (0.114) |
0.748 (0.479) |
0.570 (0.597) |
|
SD-SC |
0.821 (0.495) |
0.123 (0.934) |
0.425 (0.686) |
0.745 (0.437) |
|
SD-EF |
3.916 (0.002) |
5.457 (<0.0001) |
4.144 (0.001) |
3.533 (0.001) |
|
SD-IC |
.002 (1) |
0.702 (0.539) |
0.738 (0.464) |
0.532 (0.598) |
|
SC-EF |
5.194 (<0.0001) |
7.722 (<0.0001) |
4.256 (0.0002) |
2.772 (0.007) |
|
SC-IC |
0.895 (0.429) |
1.018 (0.343) |
1.210 (0.272) |
1.172 (0.267) |
|
EF-IC |
6.543 (0.0002) |
4.272 (<0.0001) |
5.675 (0.0002) |
4.271 (0.0009) |
Cuadro S4. Valores de R de las comparaciones por pares de la fauna analizada en el Farallón y sitios de la costa occidental del golfo de California
Table S4. R values for pairwise comparisons of fauna analyzed at Farallón and sites on the western coast of the Gulf of California
|
Prueba |
Fauna total |
Peces |
Asteroideos |
Equinoideos |
|
EC-SD |
0.16 |
0.10 |
0.69 |
0.18 |
|
EC-SC |
0.48 |
0.37 |
0.48 |
0.10 |
|
EC-IC |
0.57 |
0.50 |
0.83 |
0.27 |
|
EC-IF |
0.81 |
0.75 |
0.95 |
0.36 |
|
SD-SC |
-0.03 |
-0.03 |
0.115 |
0.03 |
|
SD-IC |
0.27 |
0.27 |
0.721 |
0.23 |
|
SD-EF |
0.57 |
0.57 |
0.40 |
0.31 |
|
SC-IC |
0.26 |
0.23 |
0.61 |
0.05 |
|
SC-EF |
0.74 |
0.68 |
0.30 |
0.32 |
|
IC-EF |
0.69 |
0.71 |
0.22 |
0.41 |
1 Universidad Autónoma de Baja California Sur, Departamento de Ciencias Marinas y Costeras. Carretera al sur km 5.5., Col. El Mezquitito, La Paz, B. C. S., 23090, México. hreyes@uabcs.mx; ORCID: https://orcid.org/0000-0003-2593-9631
2 Laboratorio de Ecología Marina, Centro de Investigaciones Costeras, Centro Universitario de la Costa, Universidad de Guadalajara. Avenida Universidad 203, Delegación Ixtapa, Puerto Vallarta, Jalisco, 48280, México. viomarcast@gmail.com; ORCID: https://orcid.org/0000-0003-3932-4646
3 Posgrado en Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México. Avenida Universidad 3000, Ciudad Universitaria Coyoacán, C. P., 04510, Ciudad de México, México. nelo.olan@gmail.com; ORCID: https://orcid.org/0000-0003-2997-8937
4 Área de Protección de Flora y Fauna Balandra, Comisión Nacional de Áreas Naturales Protegidas. Melchor Ocampo #1045, Col. Centro, 23000, La Paz, B. C. S., México. barbara.rojas89@gmail.com, barbara.rojas@undp.org; ORCID: https://orcid.org/0000-0002-1533-499X
|
|




